

Exosites in Hypervariable Loops of ADAMTS Spacer Domains control Substrate Recognition and Proteolysis
- PMID: 31358852
- PMCID: PMC6662762
- DOI: 10.1038/s41598-019-47494-w
Exosites in Hypervariable Loops of ADAMTS Spacer Domains control Substrate Recognition and Proteolysis
Abstract
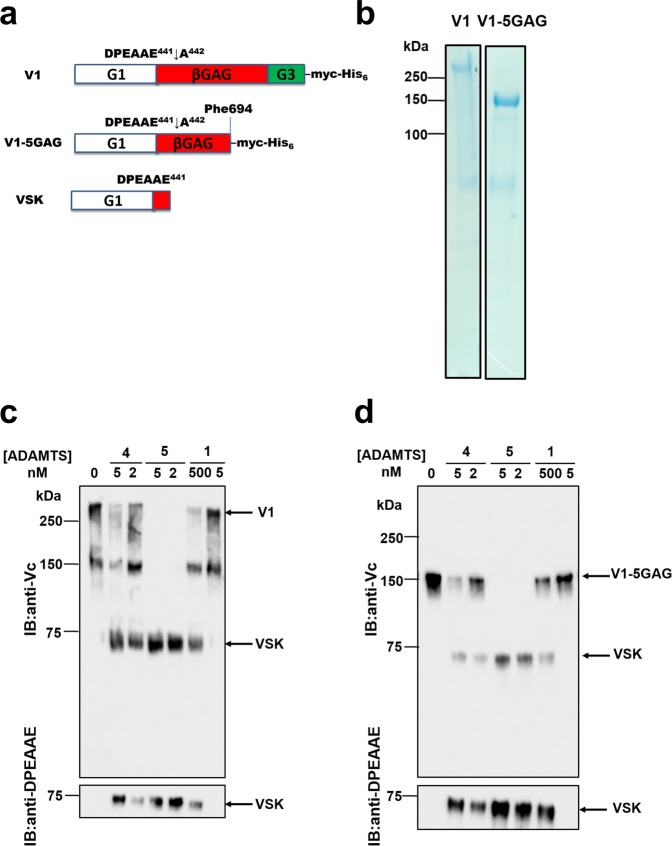
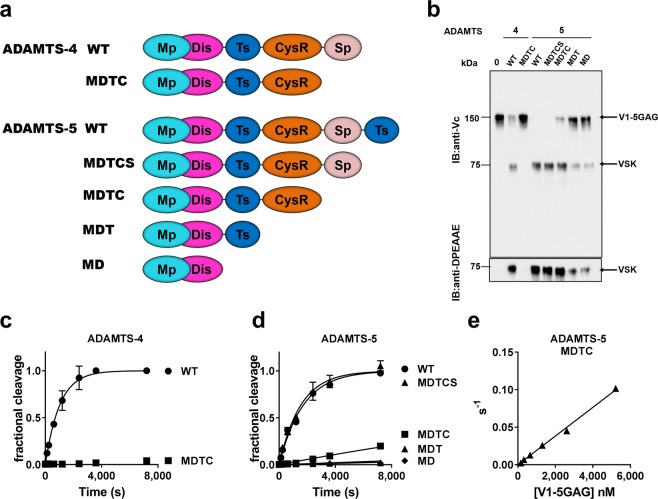
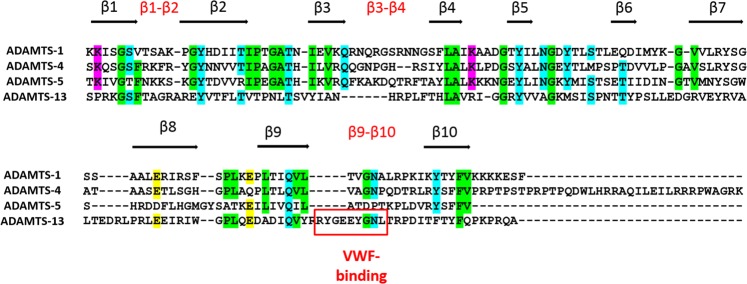
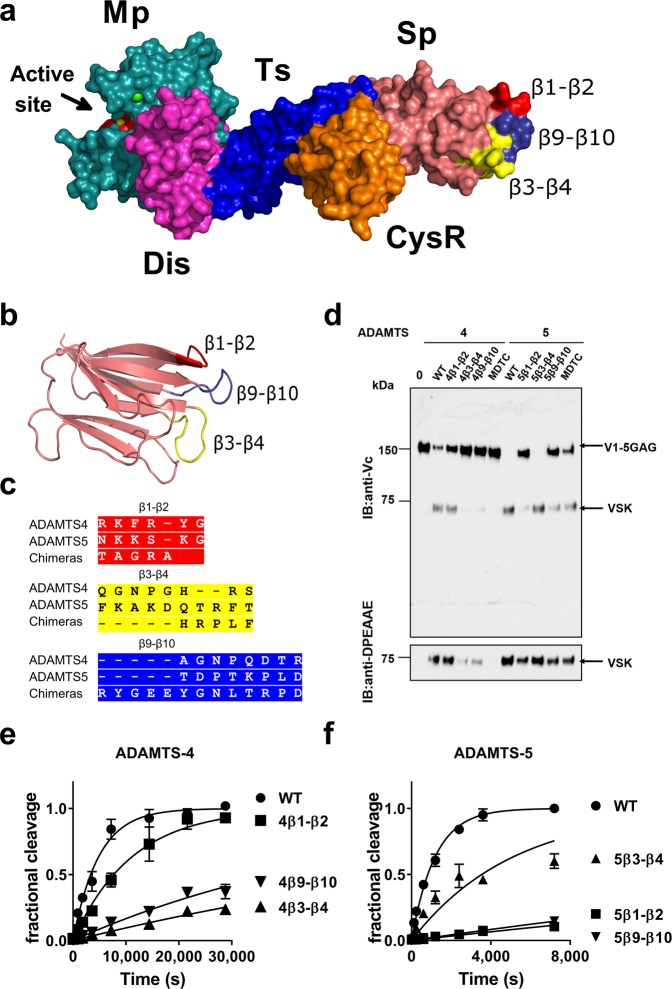
ADAMTS (A Disintegrin-like and Metalloproteinase domain with Thrombospondin type 1 Motif)-1, -4 and -5 share the abilities to cleave large aggregating proteoglycans including versican and aggrecan. These activities are highly relevant to cardiovascular disease and osteoarthritis and during development. Here, using purified recombinant ADAMTS-1, -4 and -5, we quantify, compare, and define the molecular basis of their versicanase activity. A novel sandwich-ELISA detecting the major versican cleavage fragment was used to determine, for the first time, kinetic constants for versican proteolysis. ADAMTS-5 (kcat/Km 35 × 105 M-1 s-1) is a more potent (~18-fold) versicanase than ADAMTS-4 (kcat/Km 1.86 × 105 M-1 sec-1), whereas ADAMTS-1 versicanase activity is comparatively low. Deletion of the spacer domain reduced versicanase activity of ADAMTS-5 19-fold and that of ADAMTS-4 167-fold. Co-deletion of the ADAMTS-5 cysteine-rich domain further reduced versicanase activity to a total 153-fold reduction. Substitution of two hypervariable loops in the spacer domain of ADAMTS-5 (residues 739-744 and 837-844) and ADAMTS-4 (residues 717-724 and 788-795) with those of ADAMTS-13, which does not cleave proteoglycans, caused spacer-dependent reductions in versicanase activities. Our results demonstrate that these loops contain exosites critical for interaction with and processing of versican. The hypervariable loops of ADAMTS-5 are shown to be important also for its aggrecanase activity. Together with previous work on ADAMTS-13 our results suggest that the spacer domain hypervariable loops may exercise significant control of ADAMTS proteolytic activity as a general principle. Identification of specific exosites also provides targets for selective inhibitors.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
