

A small molecule inhibitor of ER-to-cytosol protein dislocation exhibits anti-dengue and anti-Zika virus activity
- PMID: 31358863
- PMCID: PMC6662757
- DOI: 10.1038/s41598-019-47532-7
A small molecule inhibitor of ER-to-cytosol protein dislocation exhibits anti-dengue and anti-Zika virus activity
Abstract
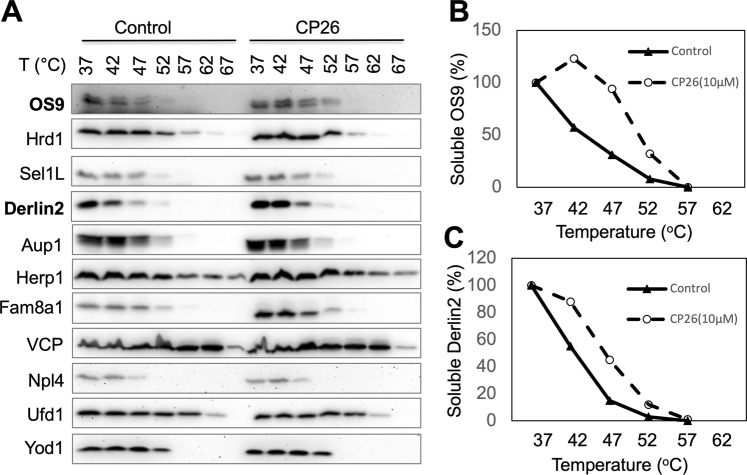
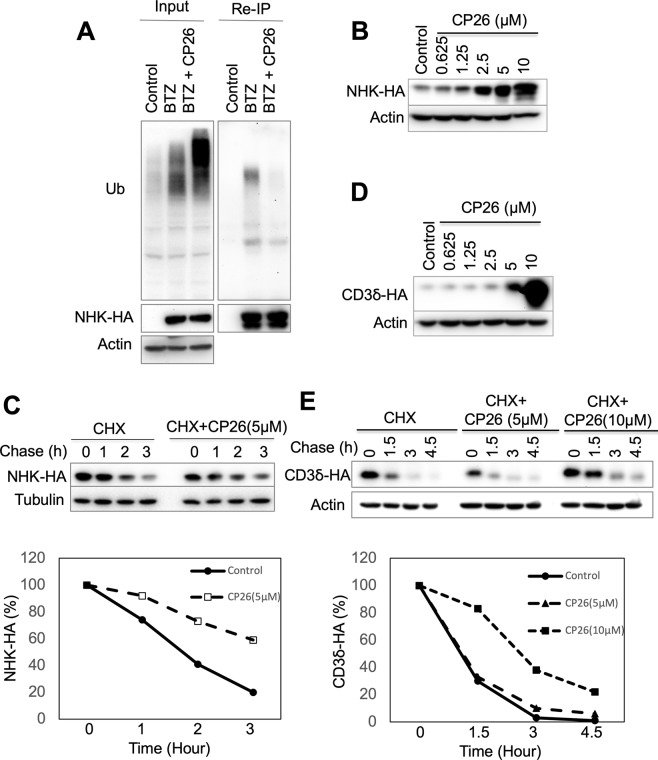
Infection with flaviviruses, such as dengue virus (DENV) and the recently re-emerging Zika virus (ZIKV), represents an increasing global risk. Targeting essential host elements required for flavivirus replication represents an attractive approach for the discovery of antiviral agents. Previous studies have identified several components of the Hrd1 ubiquitin ligase-mediated endoplasmic reticulum (ER)-associated degradation (ERAD) pathway, a cellular protein quality control process, as host factors crucial for DENV and ZIKV replication. Here, we report that CP26, a small molecule inhibitor of protein dislocation from the ER lumen to the cytosol, which is an essential step for ERAD, has broad-spectrum anti-flavivirus activity. CP26 targets the Hrd1 complex, inhibits ERAD, and induces ER stress. Ricin and cholera toxins are known to hijack the protein dislocation machinery to reach the cytosol, where they exert their cytotoxic effects. CP26 selectively inhibits the activity of cholera toxin but not that of ricin. CP26 exhibits a significant inhibitory activity against both DENV and ZIKV, providing substantial protection to the host cells against virus-induced cell death. This study identified a novel dislocation inhibitor, CP26, that shows potent anti-DENV and anti-ZIKV activity in cells. Furthermore, this study provides the first example of the targeting of host ER dislocation with small molecules to combat flavivirus infection.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
