

The Dynamic 3D Genome in Gametogenesis and Early Embryonic Development
- PMID: 31362461
- PMCID: PMC6721571
- DOI: 10.3390/cells8080788
The Dynamic 3D Genome in Gametogenesis and Early Embryonic Development
Abstract
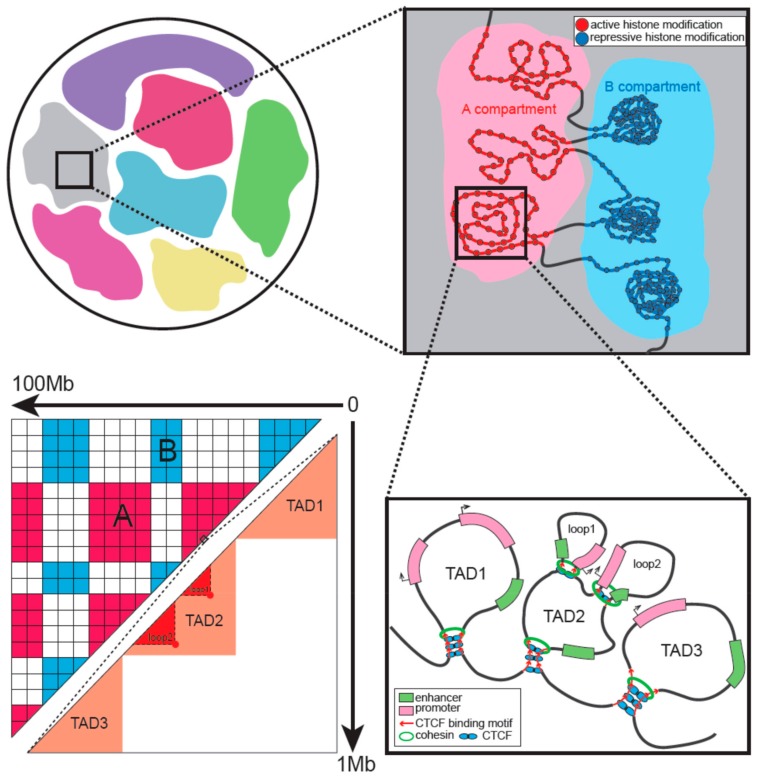
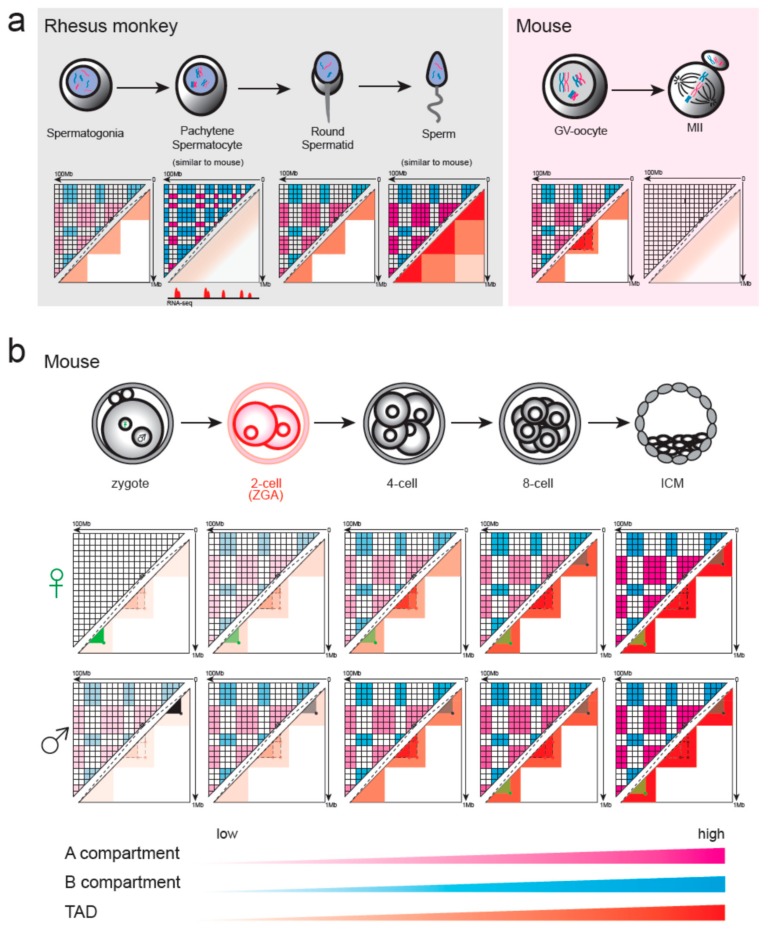
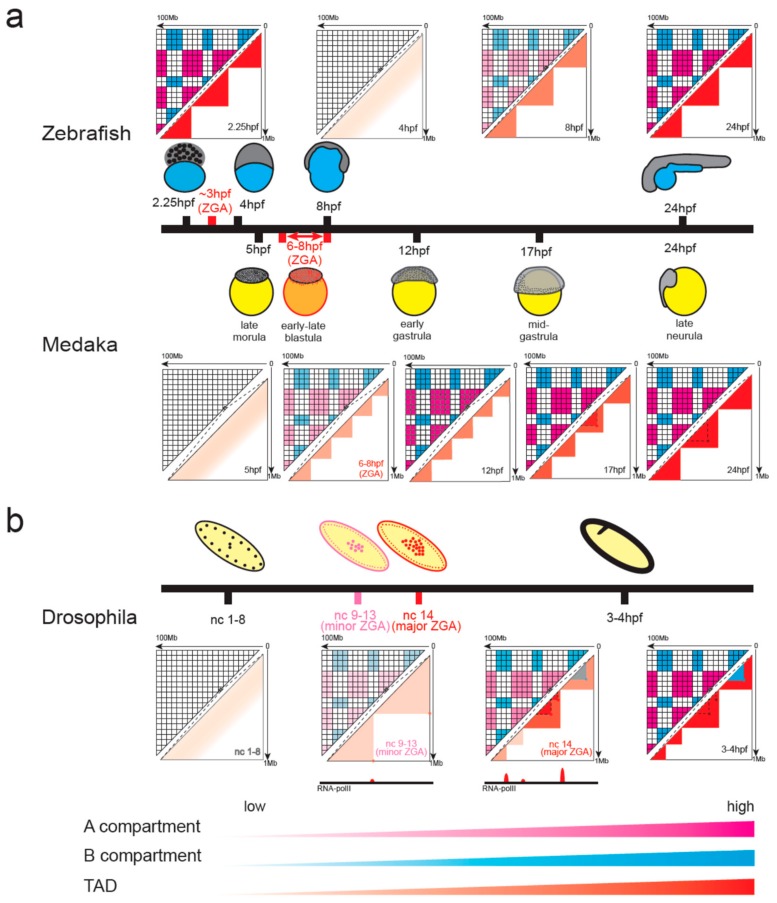
During gametogenesis and early embryonic development, the chromatin architecture changes dramatically, and both the transcriptomic and epigenomic landscape are comprehensively reprogrammed. Understanding these processes is the holy grail in developmental biology and a key step towards evolution. The 3D conformation of chromatin plays a central role in the organization and function of nuclei. Recently, the dynamics of chromatin structures have been profiled in many model and non-model systems, from insects to mammals, resulting in an interesting comparison. In this review, we first introduce the research methods of 3D chromatin structure with low-input material suitable for embryonic study. Then, the dynamics of 3D chromatin architectures during gametogenesis and early embryonic development is summarized and compared between species. Finally, we discuss the possible mechanisms for triggering the formation of genome 3D conformation in early development.
Keywords: chromatin structure; early embryonic development; formation mechanism of 3D genome; gametogenesis; low-input Hi-C; single-cell Hi-C.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
