

Pseudomonas aeruginosa Increases the Sensitivity of Biofilm-Grown Staphylococcus aureus to Membrane-Targeting Antiseptics and Antibiotics
- PMID: 31363032
- PMCID: PMC6667622
- DOI: 10.1128/mBio.01501-19
Pseudomonas aeruginosa Increases the Sensitivity of Biofilm-Grown Staphylococcus aureus to Membrane-Targeting Antiseptics and Antibiotics
Abstract
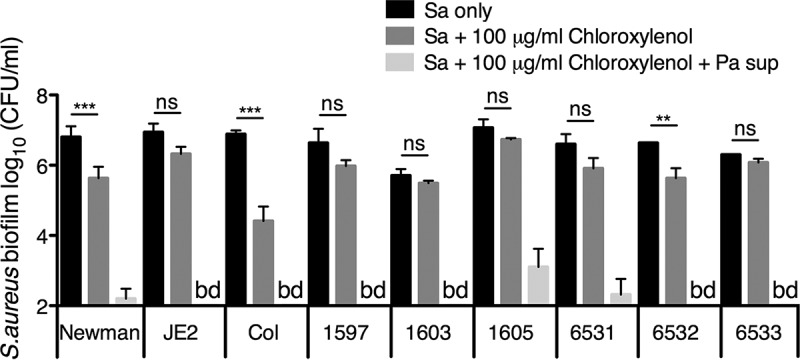
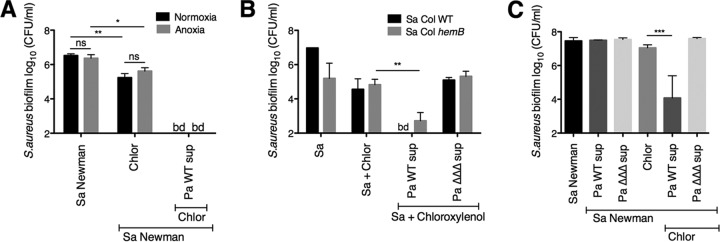
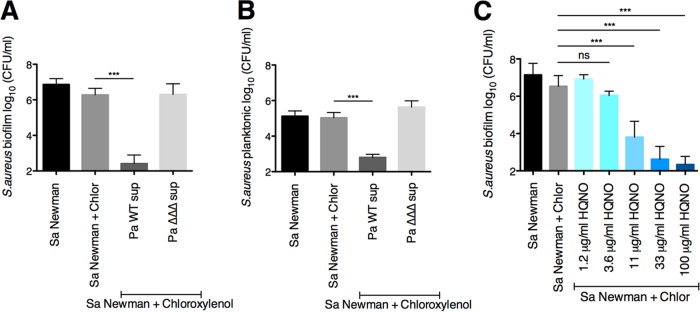
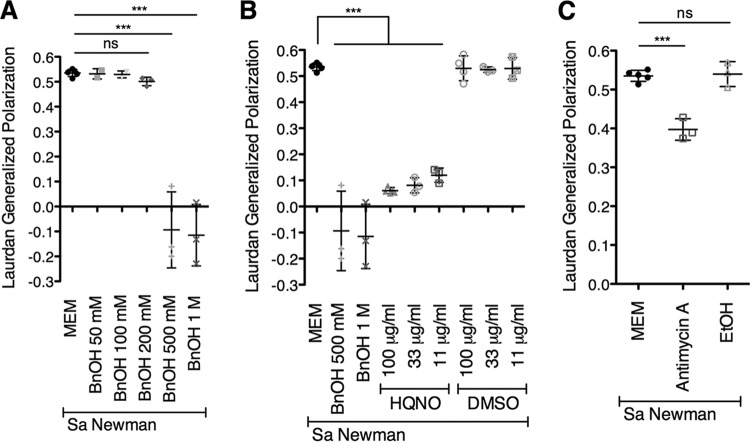
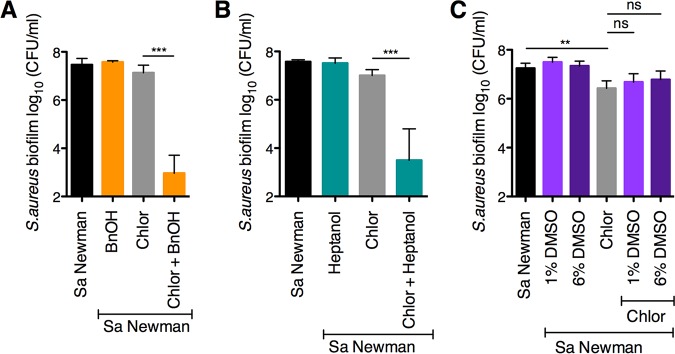
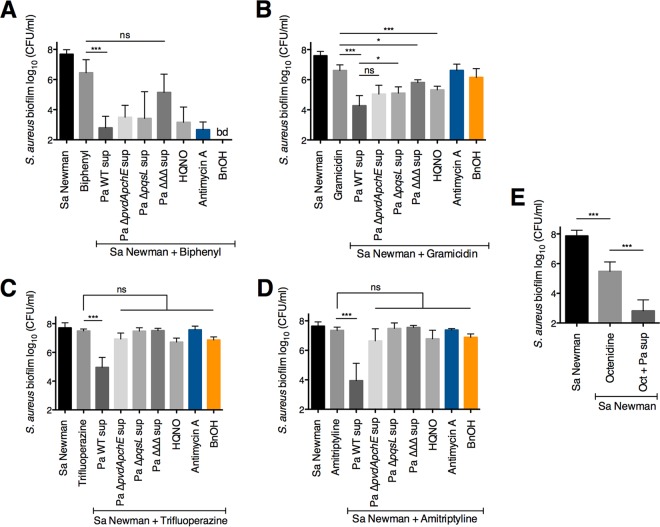
Pseudomonas aeruginosa and Staphylococcus aureus often cause chronic, recalcitrant infections in large part due to their ability to form biofilms. The biofilm mode of growth enables these organisms to withstand antibacterial insults that would effectively eliminate their planktonic counterparts. We found that P. aeruginosa supernatant increased the sensitivity of S. aureus biofilms to multiple antimicrobial compounds, including fluoroquinolones and membrane-targeting antibacterial agents, including the antiseptic chloroxylenol. Treatment of S. aureus with the antiseptic chloroxylenol alone did not decrease biofilm cell viability; however, the combination of chloroxylenol and P. aeruginosa supernatant led to a 4-log reduction in S. aureus biofilm viability compared to exposure to chloroxylenol alone. We found that the P. aeruginosa-produced small molecule 2-n-heptyl-4-hydroxyquinoline N-oxide (HQNO) is responsible for the observed heightened sensitivity of S. aureus to chloroxylenol. Similarly, HQNO increased the susceptibility of S. aureus biofilms to other compounds, including both traditional and nontraditional antibiotics, which permeabilize bacterial membranes. Genetic and phenotypic studies support a model whereby HQNO causes an increase in S. aureus membrane fluidity, thereby improving the efficacy of membrane-targeting antiseptics and antibiotics. Importantly, our data show that P. aeruginosa exoproducts can enhance the ability of various antimicrobial agents to kill biofilm populations of S. aureus that are typically difficult to eradicate. Finally, our discovery that altering membrane fluidity shifts antimicrobial sensitivity profiles of bacterial biofilms may guide new approaches to target persistent infections, such as those commonly found in respiratory tract infections and in chronic wounds.IMPORTANCE The thick mucus in the airways of cystic fibrosis (CF) patients predisposes them to frequent, polymicrobial respiratory infections. Pseudomonas aeruginosa and Staphylococcus aureus are frequently coisolated from the airways of individuals with CF, as well as from diabetic foot ulcers and other wounds. Both organisms form biofilms, which are notoriously difficult to eradicate and promote chronic infection. In this study, we have shown that P. aeruginosa-secreted factors can increase the efficacy of compounds that alone have little or no bactericidal activity against S. aureus biofilms. In particular, we discovered that P. aeruginosa exoproducts can potentiate the antistaphylococcal activity of phenol-based antiseptics and other membrane-active drugs. Our findings illustrate that polymicrobial interactions can dramatically increase antibacterial efficacy in vitro and suggest that altering membrane physiology promotes the ability of certain drugs to kill bacterial biofilms-knowledge that may provide a path for the discovery of new biofilm-targeting antimicrobial strategies.
Keywords: Pseudomonas aeruginosa; Staphylococcus aureus; antibiotics; biofilm; membrane.
Copyright © 2019 Orazi et al.
Figures
Similar articles
-
Pseudomonas aeruginosa PA14 Enhances the Efficacy of Norfloxacin against Staphylococcus aureus Newman Biofilms.J Bacteriol. 2020 Aug 25;202(18):e00159-20. doi: 10.1128/JB.00159-20. Print 2020 Aug 25. J Bacteriol. 2020. PMID: 32661077 Free PMC article.
-
Pseudomonas aeruginosa Alters Staphylococcus aureus Sensitivity to Vancomycin in a Biofilm Model of Cystic Fibrosis Infection.mBio. 2017 Jul 18;8(4):e00873-17. doi: 10.1128/mBio.00873-17. mBio. 2017. PMID: 28720732 Free PMC article.
-
A Pseudomonas aeruginosa Antimicrobial Affects the Biogeography but Not Fitness of Staphylococcus aureus during Coculture.mBio. 2021 Mar 30;12(2):e00047-21. doi: 10.1128/mBio.00047-21. mBio. 2021. PMID: 33785630 Free PMC article.
-
Pseudomonas aeruginosa and Staphylococcus aureus communication in biofilm infections: insights through network and database construction.Crit Rev Microbiol. 2019 Sep-Nov;45(5-6):712-728. doi: 10.1080/1040841X.2019.1700209. Epub 2019 Dec 13. Crit Rev Microbiol. 2019. PMID: 31835971 Review.
-
Promising treatment strategies to combat Staphylococcus aureus biofilm infections: an updated review.Biofouling. 2020 Nov;36(10):1159-1181. doi: 10.1080/08927014.2020.1857743. Epub 2020 Dec 22. Biofouling. 2020. PMID: 33353409 Review.
Cited by
-
Bidirectional alterations in antibiotics susceptibility in Staphylococcus aureus-Pseudomonas aeruginosa dual-species biofilm.Sci Rep. 2020 Sep 9;10(1):14849. doi: 10.1038/s41598-020-71834-w. Sci Rep. 2020. PMID: 32908166 Free PMC article.
-
Strain Background, Species Frequency, and Environmental Conditions Are Important in Determining Pseudomonas aeruginosa and Staphylococcus aureus Population Dynamics and Species Coexistence.Appl Environ Microbiol. 2020 Sep 1;86(18):e00962-20. doi: 10.1128/AEM.00962-20. Print 2020 Sep 1. Appl Environ Microbiol. 2020. PMID: 32651205 Free PMC article.
-
"It Takes a Village": Mechanisms Underlying Antimicrobial Recalcitrance of Polymicrobial Biofilms.J Bacteriol. 2019 Dec 6;202(1):e00530-19. doi: 10.1128/JB.00530-19. Print 2019 Dec 6. J Bacteriol. 2019. PMID: 31548277 Free PMC article. Review.
-
Supplemental Oxygen Alters the Airway Microbiome in Cystic Fibrosis.mSystems. 2022 Oct 26;7(5):e0036422. doi: 10.1128/msystems.00364-22. Epub 2022 Aug 24. mSystems. 2022. PMID: 36000724 Free PMC article.
-
Pseudomonas aeruginosa surface motility and invasion into competing communities enhances interspecies antagonism.bioRxiv [Preprint]. 2024 Apr 4:2024.04.03.588010. doi: 10.1101/2024.04.03.588010. bioRxiv. 2024. Update in: mBio. 2024 Sep 11;15(9):e0095624. doi: 10.1128/mbio.00956-24. PMID: 38617332 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
