

Modification of messenger RNA by 2'-O-methylation regulates gene expression in vivo
- PMID: 31363086
- PMCID: PMC6667457
- DOI: 10.1038/s41467-019-11375-7
Modification of messenger RNA by 2'-O-methylation regulates gene expression in vivo
Abstract
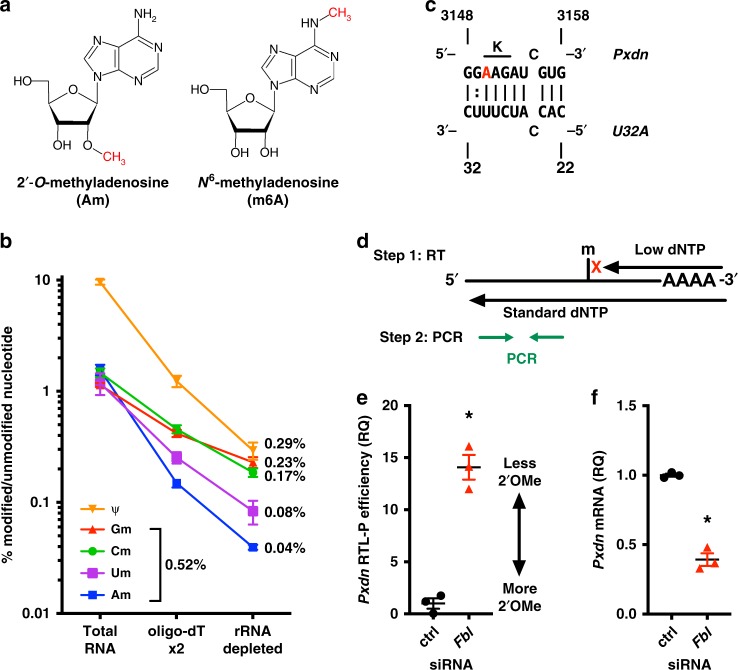
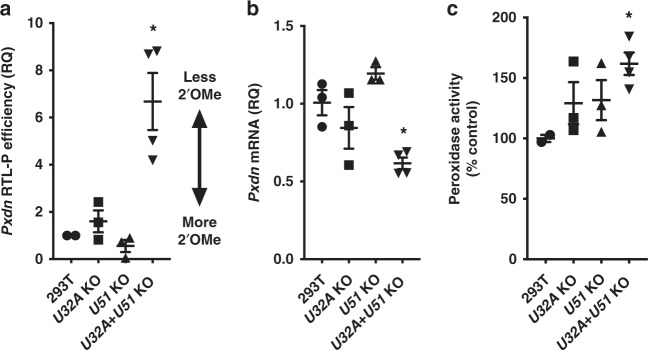
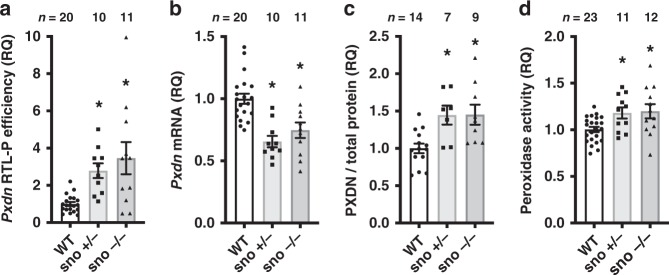
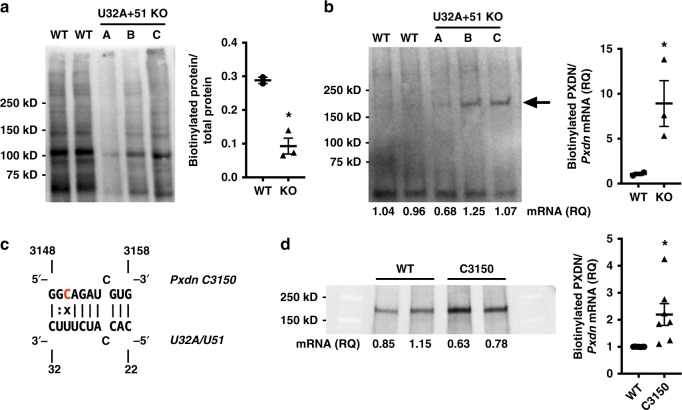
Epitranscriptomic modifications of mRNA are important regulators of gene expression. While internal 2'-O-methylation (Nm) has been discovered on mRNA, questions remain about its origin and function in cells and organisms. Here, we show that internal Nm modification can be guided by small nucleolar RNAs (snoRNAs), and that these Nm sites can regulate mRNA and protein expression. Specifically, two box C/D snoRNAs (SNORDs) and the 2'-O-methyltransferase fibrillarin lead to Nm modification in the protein-coding region of peroxidasin (Pxdn). The presence of Nm modification increases Pxdn mRNA expression but inhibits its translation, regulating PXDN protein expression and enzyme activity both in vitro and in vivo. Our findings support a model in which snoRNA-guided Nm modifications of mRNA can regulate physiologic gene expression by altering mRNA levels and tuning protein translation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
