

EZH2 cooperates with E2F1 to stimulate expression of genes involved in adrenocortical carcinoma aggressiveness
- PMID: 31363169
- PMCID: PMC6738105
- DOI: 10.1038/s41416-019-0538-y
EZH2 cooperates with E2F1 to stimulate expression of genes involved in adrenocortical carcinoma aggressiveness
Abstract
Background: EZH2 is overexpressed and associated with poor prognosis in adrenocortical carcinoma (ACC) and its inhibition reduces growth and aggressiveness of ACC cells in culture. Although EZH2 was identified as the methyltransferase that deposits the repressive H3K27me3 histone mark, it can cooperate with transcription factors to stimulate gene transcription.
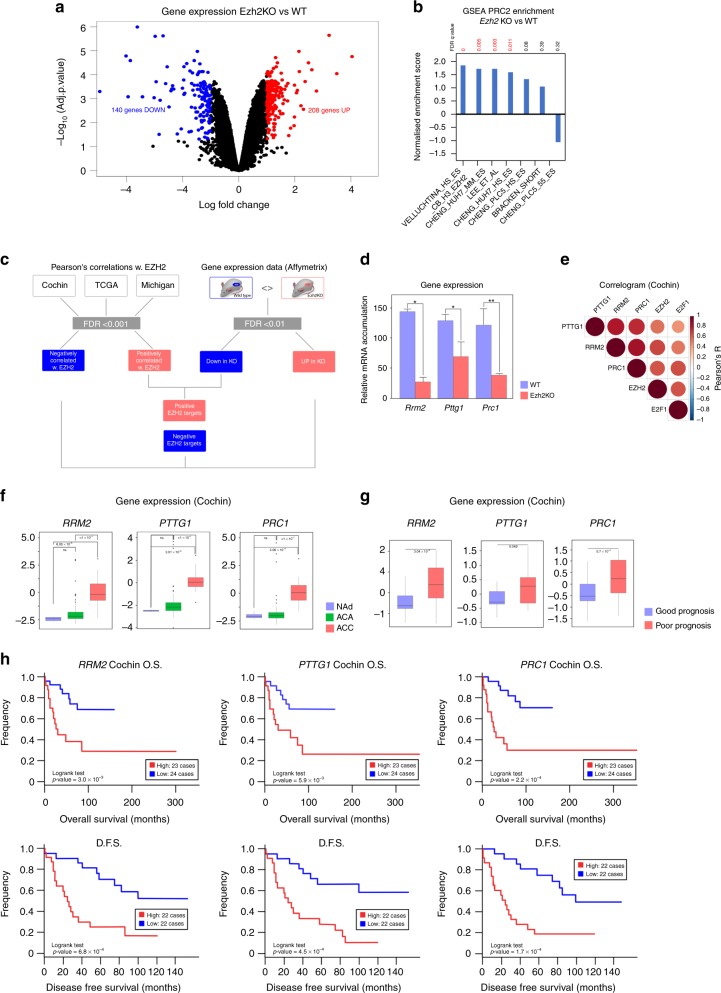
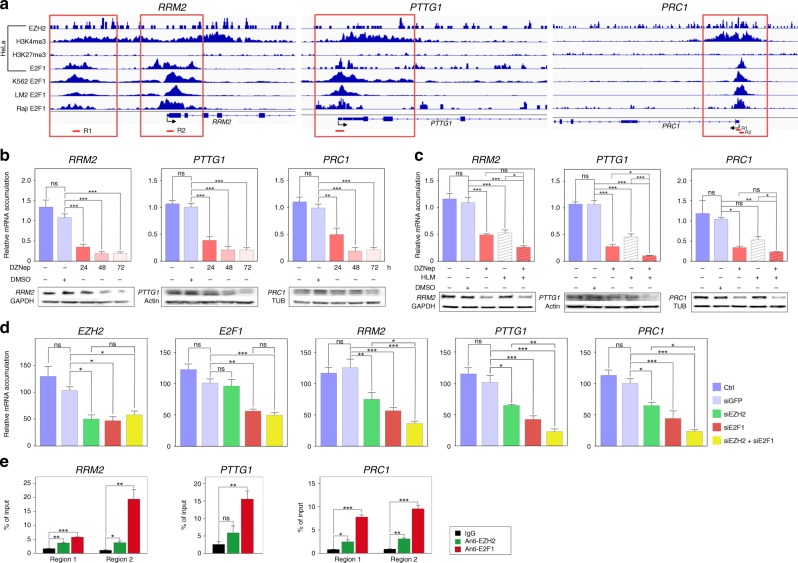
Methods: We used bioinformatics approaches on gene expression data from three cohorts of patients and a mouse model of EZH2 ablation, to identify targets and mode of action of EZH2 in ACC. This was followed by ChIP and functional assays to evaluate contribution of identified targets to ACC pathogenesis.
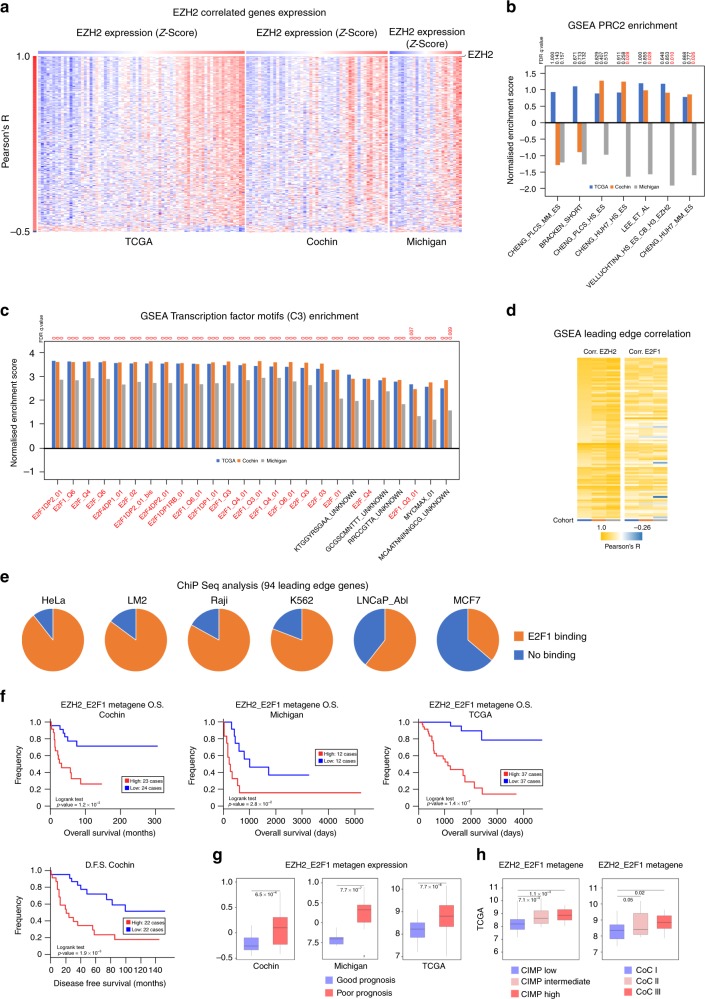
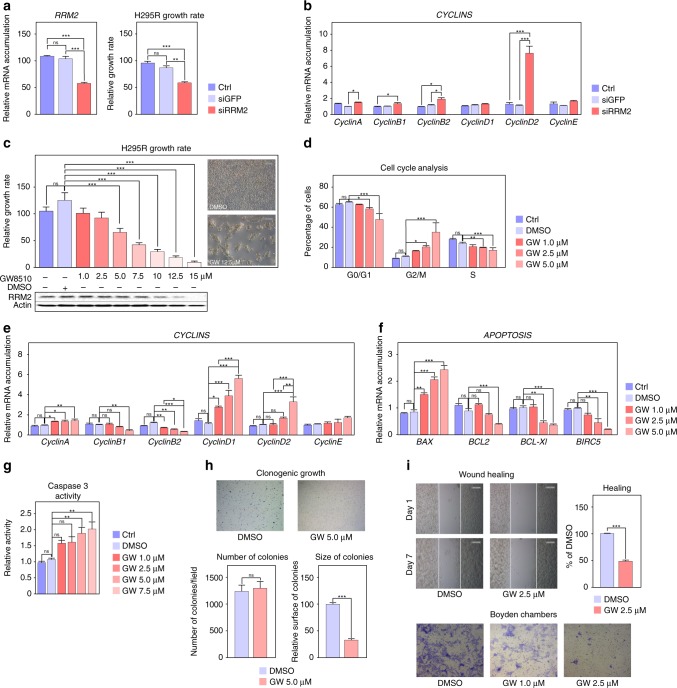
Results: We show that EZH2 mostly works as a transcriptional inducer in ACC, through cooperation with the transcription factor E2F1 and identify three positive targets involved in cell cycle regulation and mitosis i.e., RRM2, PTTG1 and ASE1/PRC1. Overexpression of these genes is associated with poor prognosis, suggesting a potential role in acquisition of aggressive ACC features. Pharmacological and siRNA-mediated inhibition of RRM2 blocks cell proliferation, induces apoptosis and inhibits cell migration, suggesting that it may be an interesting target in ACC.
Conclusions: Altogether, these data show an unexpected role of EZH2 and E2F1 in stimulating expression of genes associated with ACC aggressiveness.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
EZH2 is overexpressed in adrenocortical carcinoma and is associated with disease progression.Hum Mol Genet. 2016 Jul 1;25(13):2789-2800. doi: 10.1093/hmg/ddw136. Epub 2016 May 5. Hum Mol Genet. 2016. PMID: 27149985 Free PMC article.
-
Activation of EZH2 and SUZ12 Regulated by E2F1 Predicts the Disease Progression and Aggressive Characteristics of Bladder Cancer.Clin Cancer Res. 2015 Dec 1;21(23):5391-403. doi: 10.1158/1078-0432.CCR-14-2680. Epub 2015 Aug 12. Clin Cancer Res. 2015. PMID: 26268246
-
PTTG1 overexpression in adrenocortical cancer is associated with poor survival and represents a potential therapeutic target.Surgery. 2013 Dec;154(6):1405-16; discussion 1416. doi: 10.1016/j.surg.2013.06.058. Surgery. 2013. PMID: 24238056 Free PMC article.
-
Wnt/β-catenin activation cooperates with loss of p53 to cause adrenocortical carcinoma in mice.Oncogene. 2020 Jul;39(30):5282-5291. doi: 10.1038/s41388-020-1358-5. Epub 2020 Jun 19. Oncogene. 2020. PMID: 32561853 Free PMC article.
-
Epigenetic regulation of cancer biology and anti-tumor immunity by EZH2.Oncotarget. 2016 Dec 20;7(51):85624-85640. doi: 10.18632/oncotarget.12928. Oncotarget. 2016. PMID: 27793053 Free PMC article. Review.
Cited by
-
Sevoflurane Offers Neuroprotection in a Cerebral Ischemia/Reperfusion Injury Rat Model Through the E2F1/EZH2/TIMP2 Regulatory Axis.Mol Neurobiol. 2022 Apr;59(4):2219-2231. doi: 10.1007/s12035-021-02602-8. Epub 2022 Jan 22. Mol Neurobiol. 2022. PMID: 35064540
-
The noncanonical role of EZH2 in cancer.Cancer Sci. 2021 Apr;112(4):1376-1382. doi: 10.1111/cas.14840. Epub 2021 Feb 24. Cancer Sci. 2021. PMID: 33615636 Free PMC article. Review.
-
PCLAF promotes neuroblastoma G1/S cell cycle progression via the E2F1/PTTG1 axis.Cell Death Dis. 2022 Feb 24;13(2):178. doi: 10.1038/s41419-022-04635-w. Cell Death Dis. 2022. PMID: 35210406 Free PMC article.
-
Screening and Validation of Significant Genes with Poor Prognosis in Pathologic Stage-I Lung Adenocarcinoma.J Oncol. 2022 Apr 11;2022:3794021. doi: 10.1155/2022/3794021. eCollection 2022. J Oncol. 2022. PMID: 35444699 Free PMC article.
-
β-Catenin-Driven Differentiation Is a Tissue-Specific Epigenetic Vulnerability in Adrenal Cancer.Cancer Res. 2023 Jul 5;83(13):2123-2141. doi: 10.1158/0008-5472.CAN-22-2712. Cancer Res. 2023. PMID: 37129912 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
