

Role of the Carboxylate in Enzyme-Catalyzed Decarboxylation of Orotidine 5'-Monophosphate: Transition State Stabilization Dominates Over Ground State Destabilization
- PMID: 31365243
- PMCID: PMC6735427
- DOI: 10.1021/jacs.9b04823
Role of the Carboxylate in Enzyme-Catalyzed Decarboxylation of Orotidine 5'-Monophosphate: Transition State Stabilization Dominates Over Ground State Destabilization
Abstract
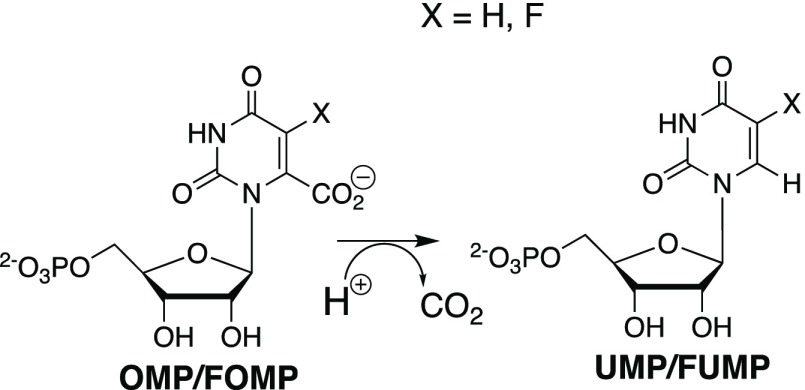
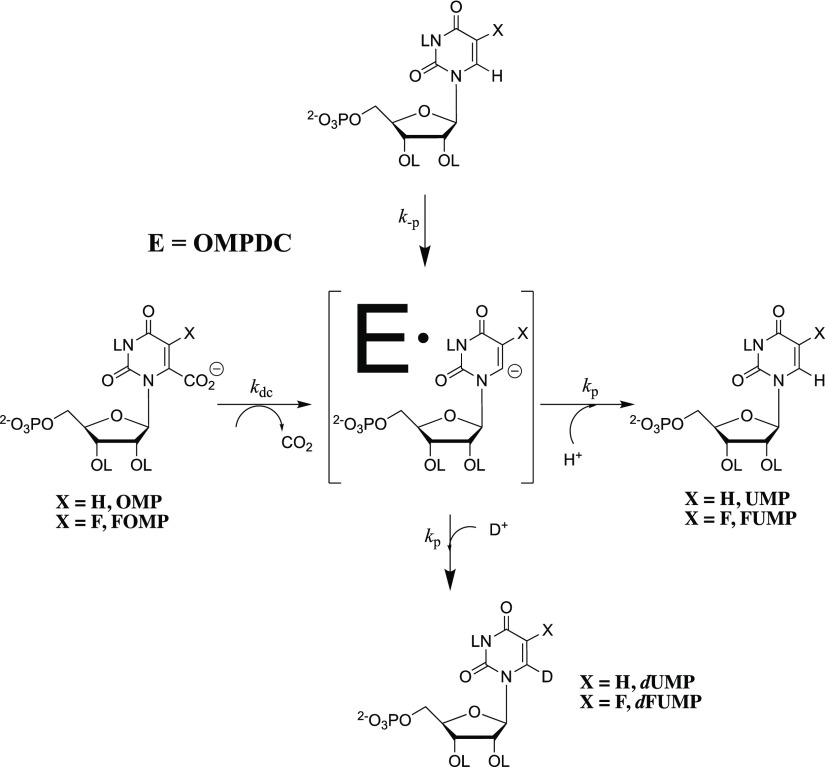
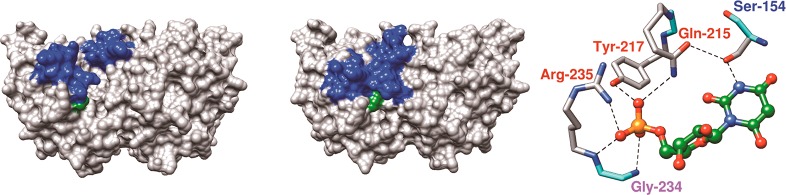
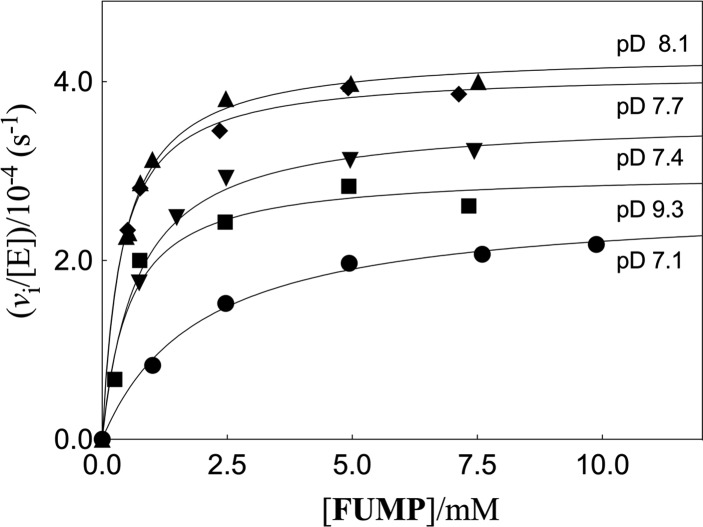
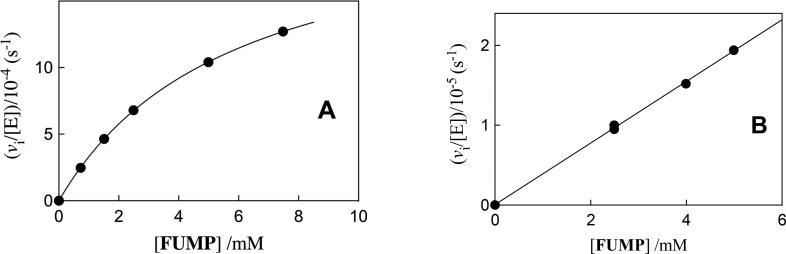
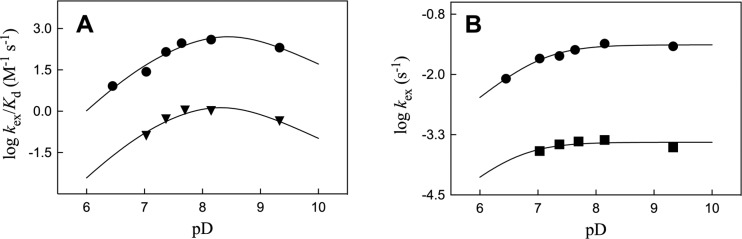
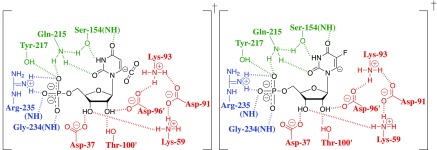
Kinetic parameters kex (s-1) and kex/Kd (M-1 s-1) are reported for exchange for deuterium in D2O of the C-6 hydrogen of 5-fluororotidine 5'-monophosphate (FUMP) catalyzed by the Q215A, Y217F, and Q215A/Y217F variants of yeast orotidine 5'-monophosphate decarboxylase (ScOMPDC) at pD 8.1, and by the Q215A variant at pD 7.1-9.3. The pD rate profiles for wildtype ScOMPDC and the Q215A variant are identical, except for a 2.5 log unit downward displacement in the profile for the Q215A variant. The Q215A, Y217F and Q215A/Y217F substitutions cause 1.3-2.0 kcal/mol larger increases in the activation barrier for wildtype ScOMPDC-catalyzed deuterium exchange compared with decarboxylation, because of the stronger apparent side chain interaction with the transition state for the deuterium exchange reaction. The stabilization of the transition state for the OMPDC-catalyzed deuterium exchange reaction of FUMP is ca. 19 kcal/mol smaller than the transition state for decarboxylation of OMP, and ca. 8 kcal/mol smaller than for OMPDC-catalyzed deprotonation of FUMP to form the vinyl carbanion intermediate common to OMPDC-catalyzed reactions OMP/FOMP and UMP/FUMP. We propose that ScOMPDC shows similar stabilizing interactions with the common portions of decarboxylation and deprotonation transition states that lead to formation of this vinyl carbanion intermediate, and that there is a large ca. (19-8) = 11 kcal/mol stabilization of the former transition state from interactions with the nascent CO2 of product. The effects of Q215A and Y217F substitutions on kcat/Km for decarboxylation of OMP are expressed mainly as an increase in Km for the reactions catalyzed by the variant enzymes, while the effects on kex/Kd for deuterium exchange are expressed mainly as an increase in kex. This shows that the Q215 and Y217 side chains stabilize the Michaelis complex to OMP for the decarboxylation reaction, compared with the complex to FUMP for the deuterium exchange reaction. These results provide strong support for the conclusion that interactions which stabilize the transition state for ScOMPDC-catalyzed decarboxylation at a nonpolar enzyme active site dominate over interactions that destabilize the ground-state Michaelis complex.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
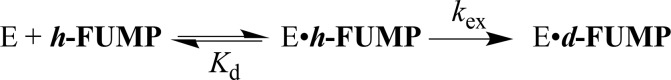
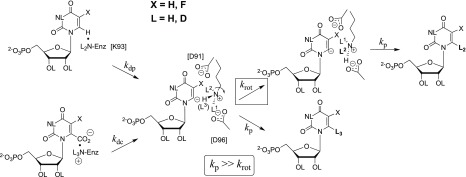
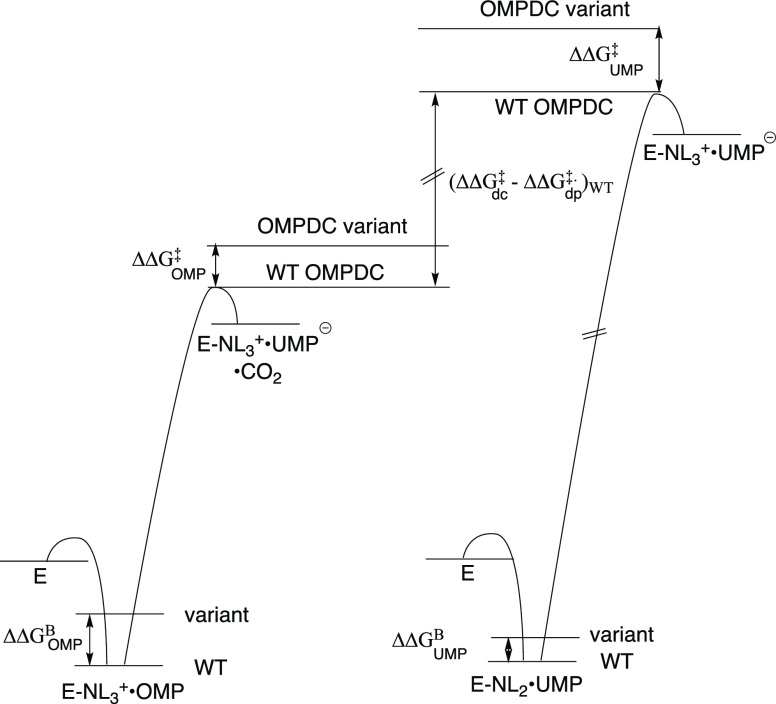
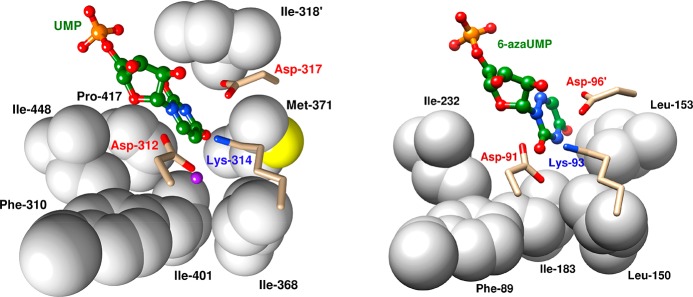
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
