

Physiology and Distribution of Archaeal Methanotrophs That Couple Anaerobic Oxidation of Methane with Sulfate Reduction
- PMID: 31366606
- PMCID: PMC6710461
- DOI: 10.1128/MMBR.00074-18
Physiology and Distribution of Archaeal Methanotrophs That Couple Anaerobic Oxidation of Methane with Sulfate Reduction
Abstract
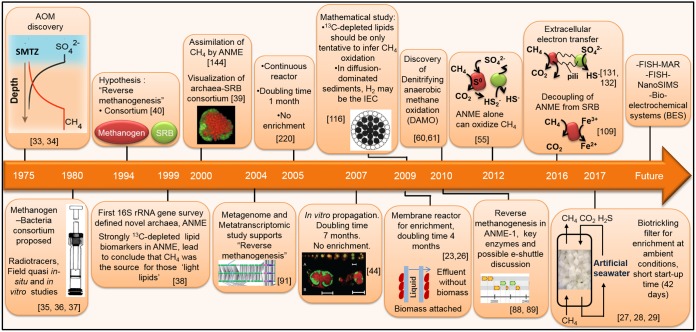
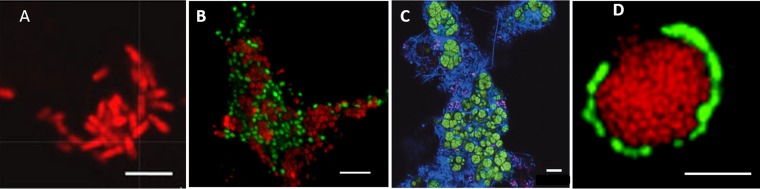
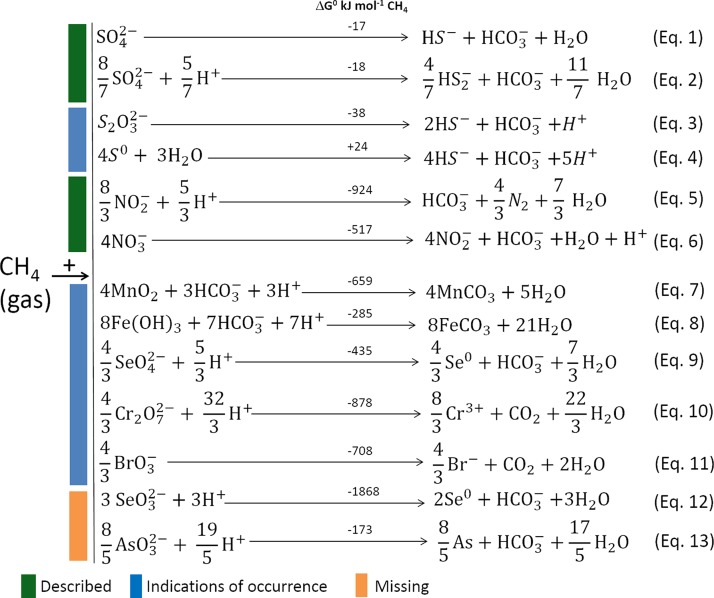
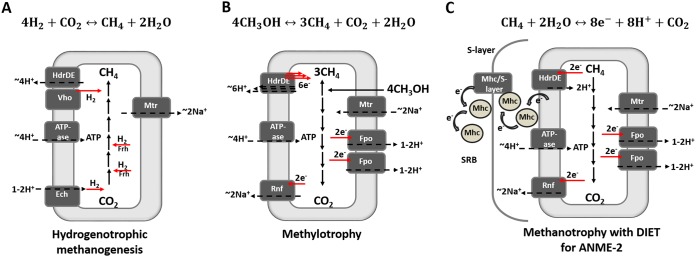
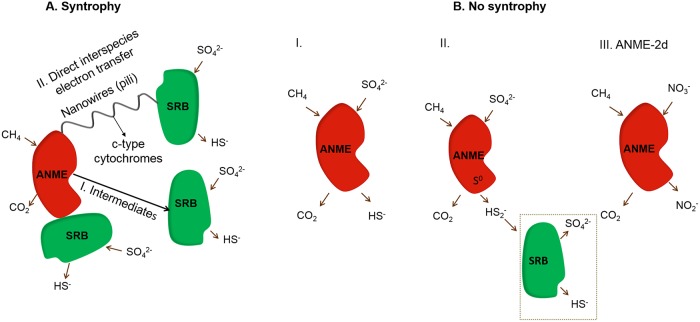
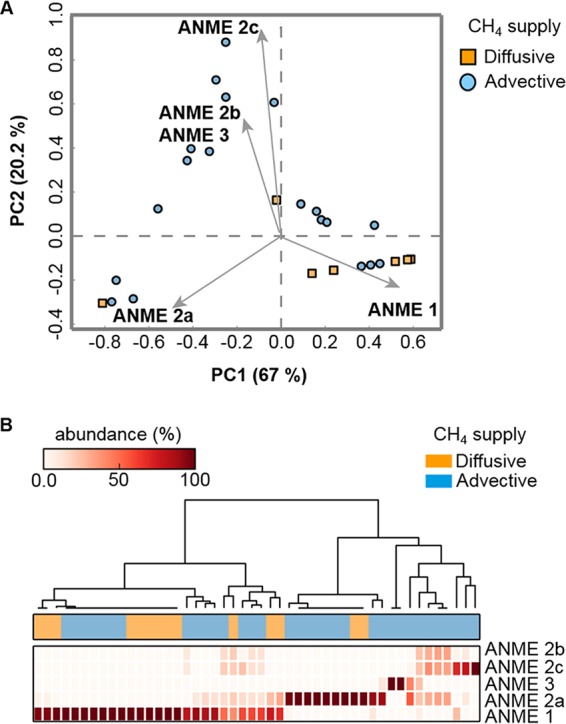
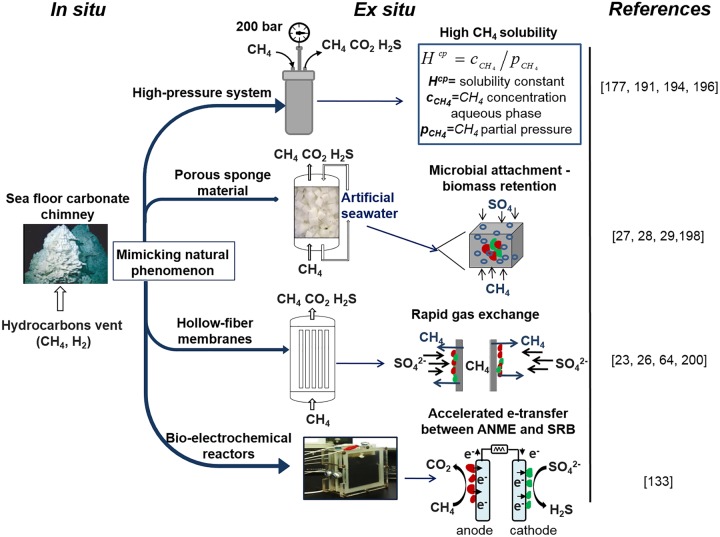
In marine anaerobic environments, methane is oxidized where sulfate-rich seawater meets biogenic or thermogenic methane. In those niches, a few phylogenetically distinct microbial types, i.e., anaerobic methanotrophs (ANME), are able to grow through anaerobic oxidation of methane (AOM). Due to the relevance of methane in the global carbon cycle, ANME have drawn the attention of a broad scientific community for 4 decades. This review presents and discusses the microbiology and physiology of ANME up to the recent discoveries, revealing novel physiological types of anaerobic methane oxidizers which challenge the view of obligate syntrophy for AOM. An overview of the drivers shaping the distribution of ANME in different marine habitats, from cold seep sediments to hydrothermal vents, is given. Multivariate analyses of the abundance of ANME in various habitats identify a distribution of distinct ANME types driven by the mode of methane transport. Intriguingly, ANME have not yet been cultivated in pure culture, despite intense attempts. Further advances in understanding this microbial process are hampered by insufficient amounts of enriched cultures. This review discusses the advantages, limitations, and potential improvements for ANME laboratory-based cultivation systems.
Keywords: anaerobic oxidation of methane; anerobic methanotrophs.
Copyright © 2019 American Society for Microbiology.
Figures

References
-
- Ciais P, Sabine C, Bala G, Bopp L, Brovkin V, Canadell J, Chhabra A, DeFries R, Galloway J, Heimann M. 2014. Carbon and other biogeochemical cycles, p 465–570. In IPCC 2013: climate change 2013. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, England.
-
- Kirschke S, Bousquet P, Ciais P, Saunois M, Canadell JG, Dlugokencky EJ, Bergamaschi P, Bergmann D, Blake DR, Bruhwiler L, Cameron-Smith P, Castaldi S, Chevallier F, Feng L, Fraser A, Heimann M, Hodson EL, Houweling S, Josse B, Fraser PJ, Krummel PB, Lamarque J-F, Langenfelds RL, Le Quere C, Naik V, O'Doherty S, Palmer PI, Pison I, Plummer D, Poulter B, Prinn RG, Rigby M, Ringeval B, Santini M, Schmidt M, Shindell DT, Simpson IJ, Spahni R, Steele LP, Strode SA, Sudo K, Szopa S, van der Werf GR, Voulgarakis A, van Weele M, Weiss RF, Williams JE, Zeng G. 2013. Three decades of global methane sources and sinks. Nat Geosci 6:813–823. doi: 10.1038/ngeo1955. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
