

Sulcal organization in the medial frontal cortex provides insights into primate brain evolution
- PMID: 31366944
- PMCID: PMC6668397
- DOI: 10.1038/s41467-019-11347-x
Sulcal organization in the medial frontal cortex provides insights into primate brain evolution
Erratum in
-
Author Correction: Sulcal organization in the medial frontal cortex provides insights into primate brain evolution.Nat Commun. 2020 Aug 6;11(1):4021. doi: 10.1038/s41467-020-17973-0. Nat Commun. 2020. PMID: 32764615 Free PMC article.
Abstract
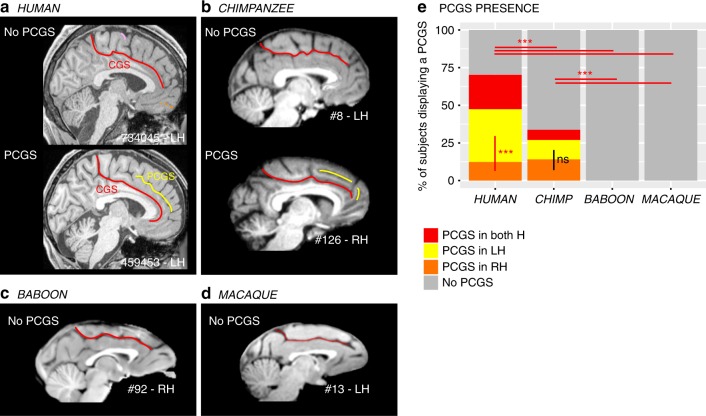
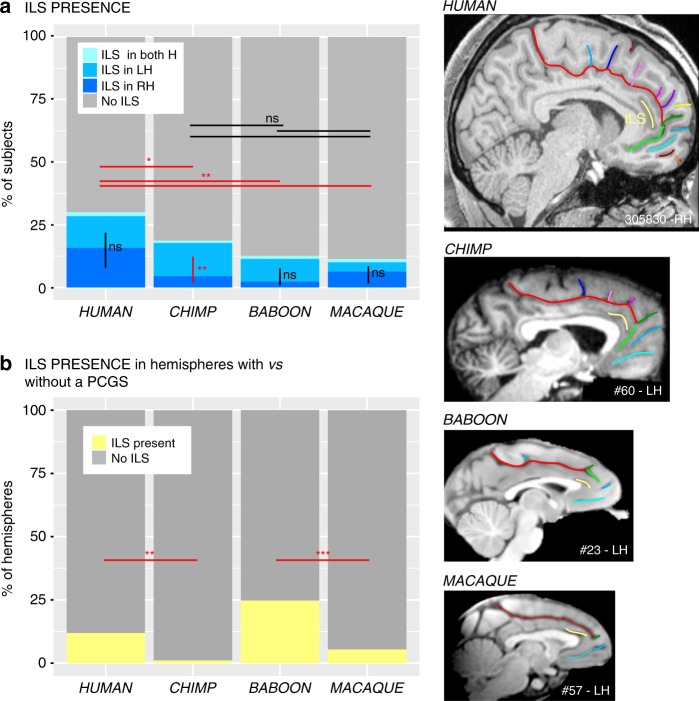
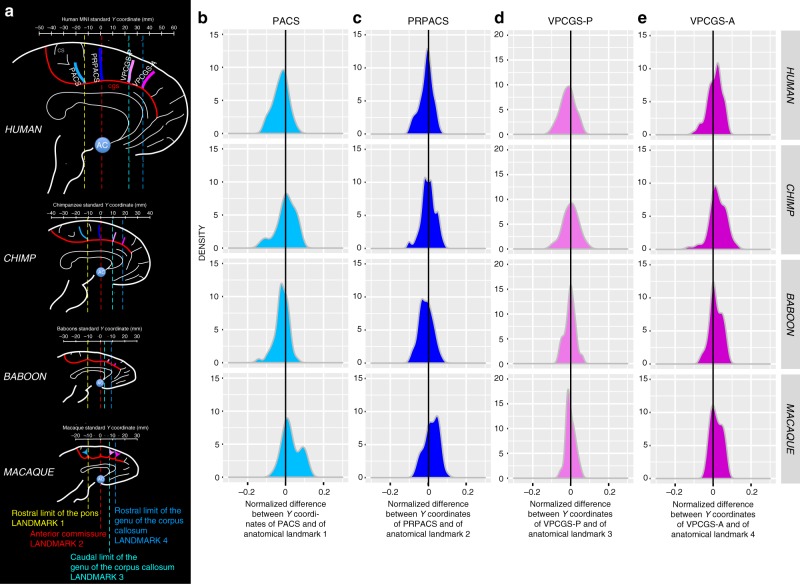
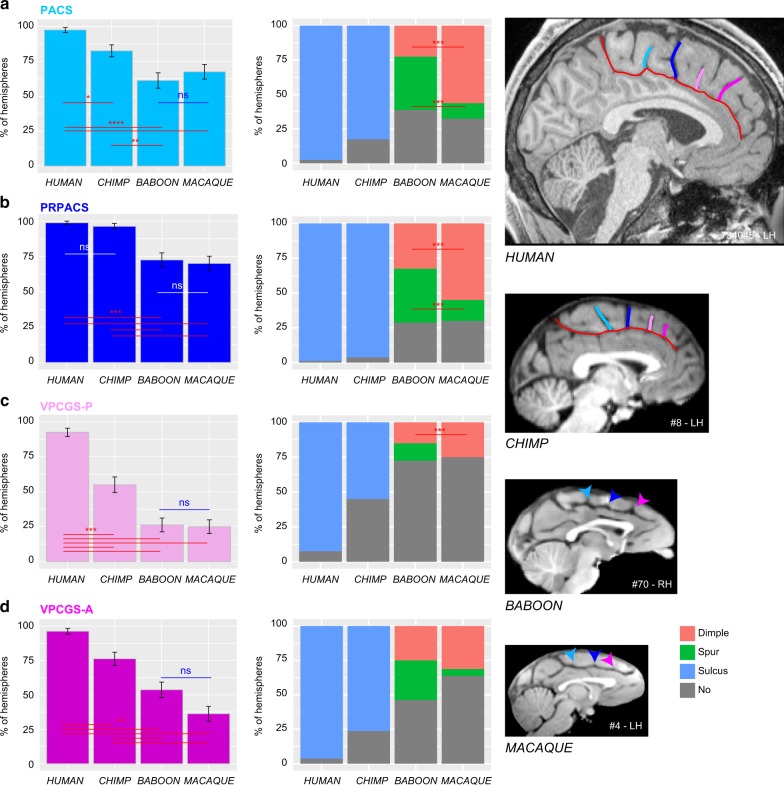
Although the relative expansion of the frontal cortex in primate evolution is generally accepted, the nature of the human uniqueness, if any, and between-species anatomo-functional comparisons of the frontal areas remain controversial. To provide a novel interpretation of the evolution of primate brains, sulcal morphological variability of the medial frontal cortex was assessed in Old World monkeys (macaque/baboon) and Hominoidea (chimpanzee/human). We show that both Hominoidea possess a paracingulate sulcus, which was previously thought to be unique to the human brain and linked to higher cognitive functions, such as mentalizing. Also, we show systematic sulcal morphological organization of the medial frontal cortex that can be traced from Old World monkeys to Hominoidea species, demonstrating an evolutionarily conserved organizational principle. These data provide a new framework to compare sulcal morphology, cytoarchitectonic areal distribution, connectivity, and function across the primate order, leading to clear predictions about how other primate brains might be anatomo-functionally organized.
Conflict of interest statement
The authors declare no competing interests.
Figures
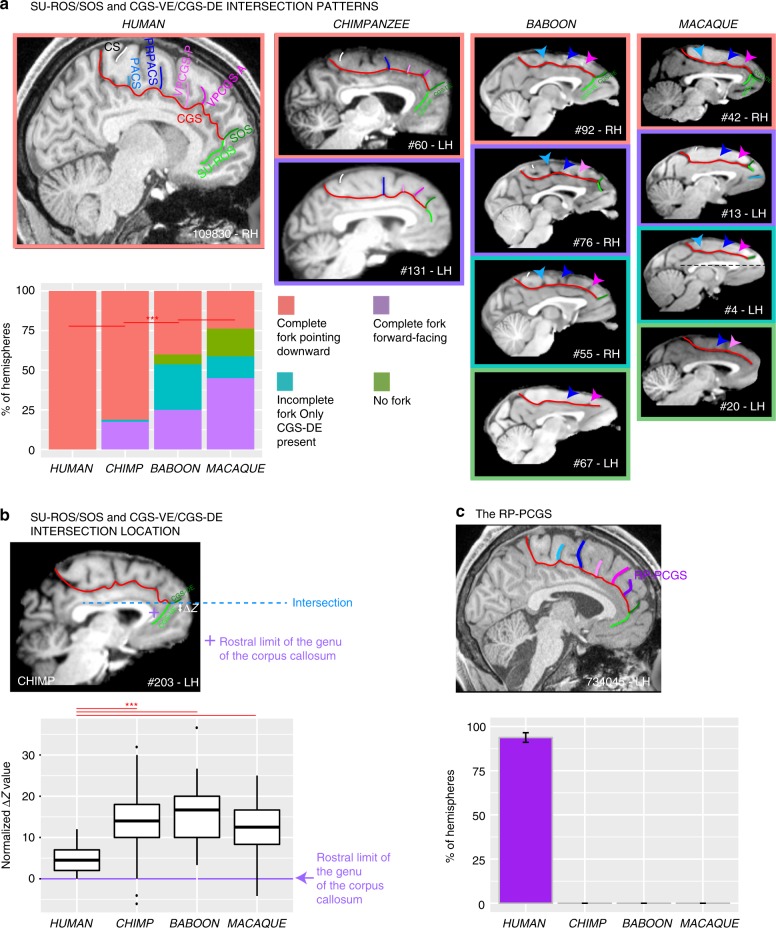
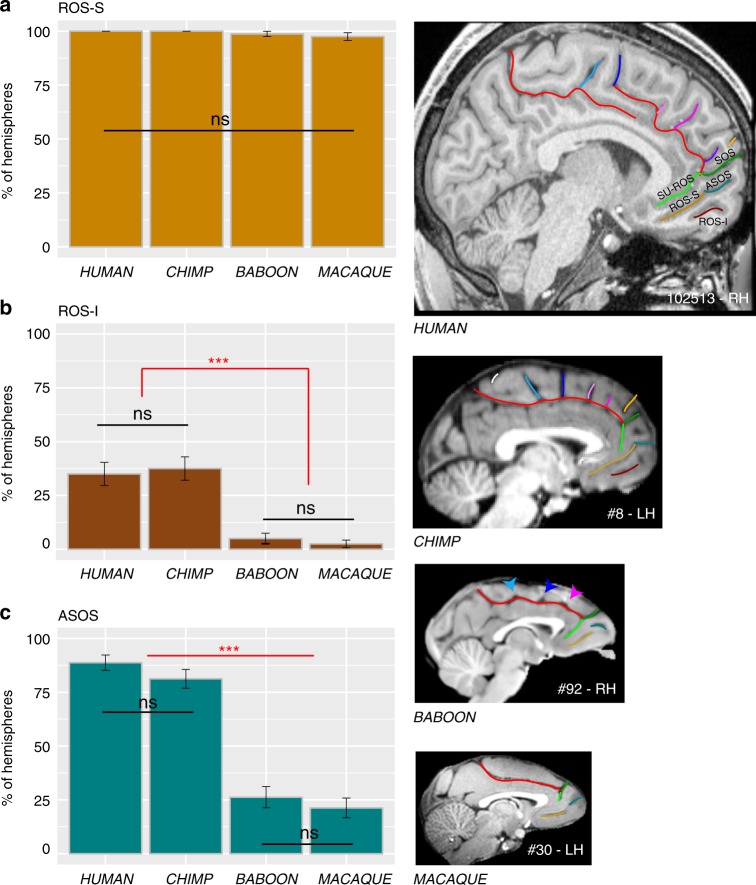
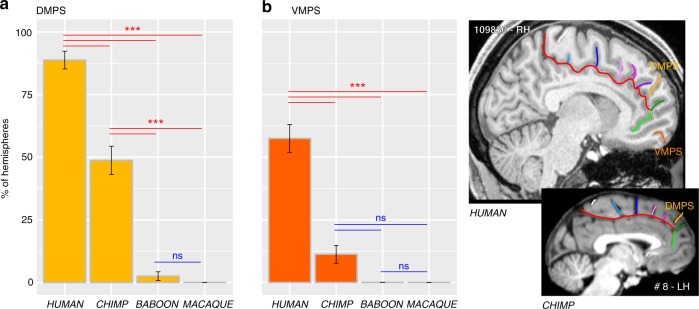
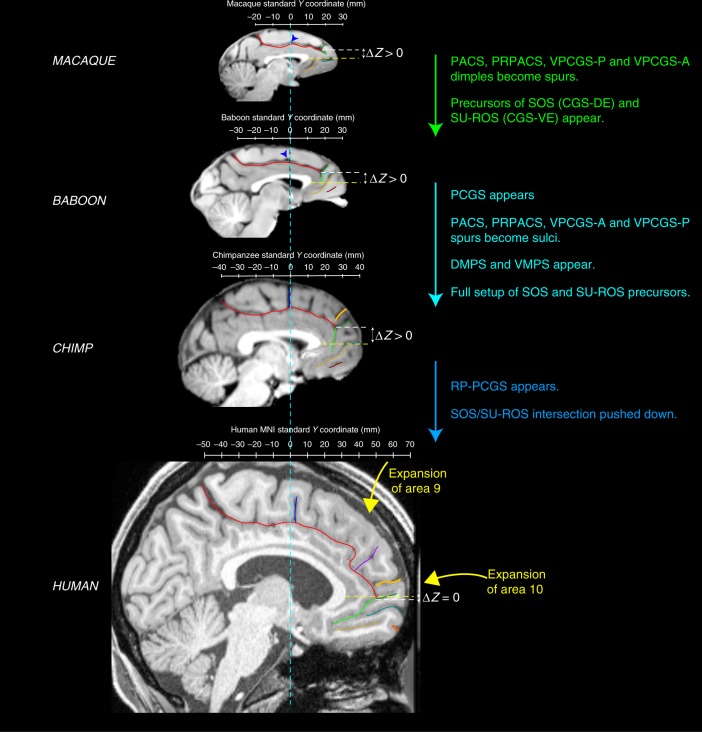
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
