

Outlook for coeliac disease patients: towards bread wheat with hypoimmunogenic gluten by gene editing of α- and γ-gliadin gene families
- PMID: 31370789
- PMCID: PMC6670228
- DOI: 10.1186/s12870-019-1889-5
Outlook for coeliac disease patients: towards bread wheat with hypoimmunogenic gluten by gene editing of α- and γ-gliadin gene families
Abstract
Background: Wheat grains contain gluten proteins, which harbour immunogenic epitopes that trigger Coeliac disease in 1-2% of the human population. Wheat varieties or accessions containing only safe gluten have not been identified and conventional breeding alone struggles to achieve such a goal, as the epitopes occur in gluten proteins encoded by five multigene families, these genes are partly located in tandem arrays, and bread wheat is allohexaploid. Gluten immunogenicity can be reduced by modification or deletion of epitopes. Mutagenesis technologies, including CRISPR/Cas9, provide a route to obtain bread wheat containing gluten proteins with fewer immunogenic epitopes.
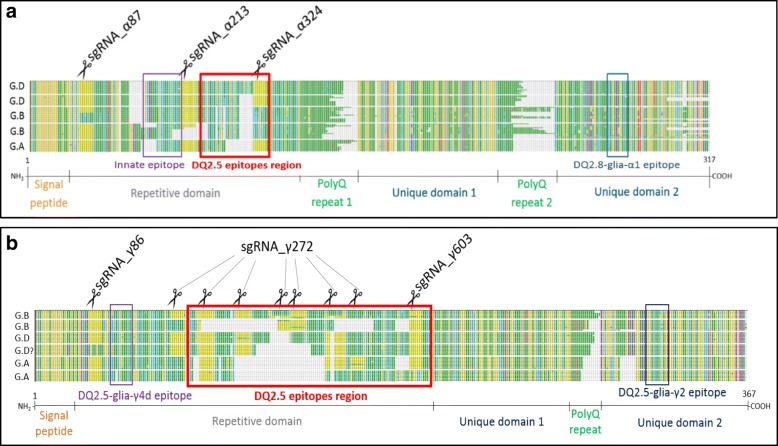
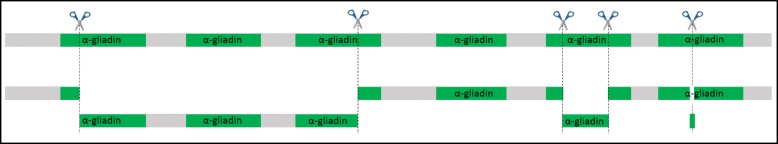
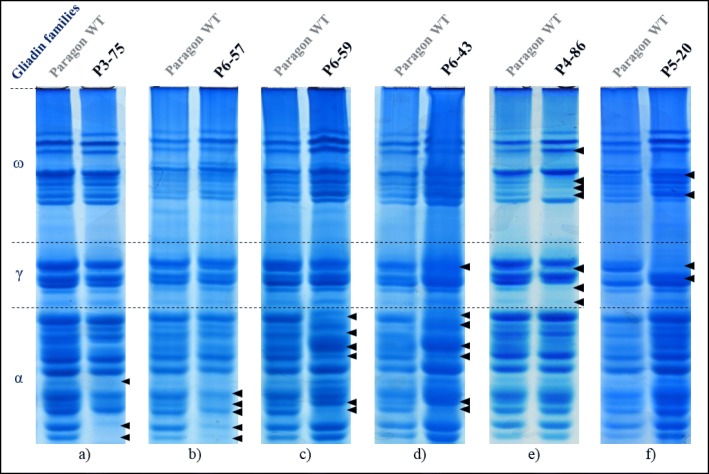
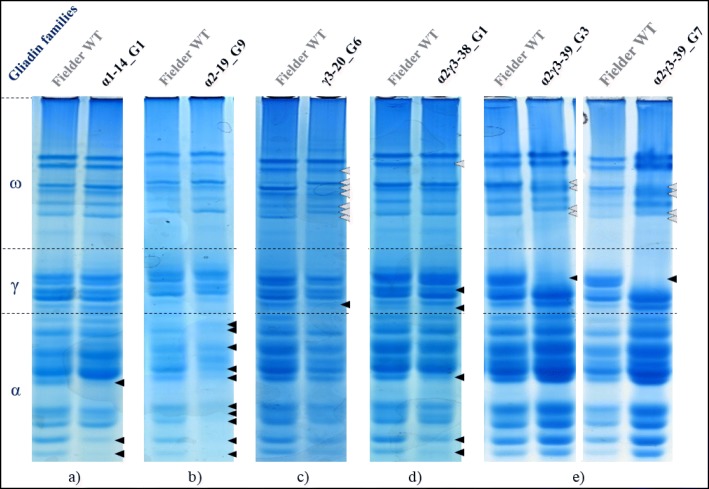
Results: In this study, we analysed the genetic diversity of over 600 α- and γ-gliadin gene sequences to design six sgRNA sequences on relatively conserved domains that we identified near coeliac disease epitopes. They were combined in four CRISPR/Cas9 constructs to target the α- or γ-gliadins, or both simultaneously, in the hexaploid bread wheat cultivar Fielder. We compared the results with those obtained with random mutagenesis in cultivar Paragon by γ-irradiation. For this, Acid-PAGE was used to identify T1 grains with altered gliadin protein profiles compared to the wild-type endosperm. We first optimised the interpretation of Acid-PAGE gels using Chinese Spring deletion lines. We then analysed the changes generated in 360 Paragon γ-irradiated lines and in 117 Fielder CRISPR/Cas9 lines. Similar gliadin profile alterations, with missing protein bands, could be observed in grains produced by both methods.
Conclusions: The results demonstrate the feasibility and efficacy of using CRISPR/Cas9 to simultaneously edit multiple genes in the large α- and γ-gliadin gene families in polyploid bread wheat. Additional methods, generating genomics and proteomics data, will be necessary to determine the exact nature of the mutations generated with both methods.
Keywords: CRISPR/Cas9; Coeliac disease; Gene editing; Gluten; Mutation breeding; Polyploid; Wheat; α-Gliadin; γ-Gliadin; γ-Irradiation.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Shewry PR, D'Ovidio R, Lafiandra D, Jenkins JA, Mills ENC, Bekes F. Wheat grain proteins. In K Khan, PR Shewry (Eds.), Wheat: Chemistry and Technology (4th edn.) St. Paul: A.A.C.C; 2009. pp. 223–298.
-
- Gilissen LJWJ, Van der Meer IM, Smulders MJM. Reducing the incidence of allergy and intolerance to cereals. J Cereal Sci. 2014;59:337–353. doi: 10.1016/j.jcs.2014.01.005. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
