

doi: 10.26508/lsa.201900408.
Print 2019 Aug.
Subunit interactions and arrangements in the fission yeast Mis16-Mis18-Mis19 complex
Affiliations
- PMID: 31371524
- PMCID: PMC6677171
- DOI: 10.26508/lsa.201900408
Item in Clipboard
Subunit interactions and arrangements in the fission yeast Mis16-Mis18-Mis19 complex
Life Sci Alliance.
.
Abstract
Centromeric chromatin in fission yeast is distinguished by the presence of nucleosomes containing the histone H3 variant Cnp1CENP-A Cell cycle-specific deposition of Cnp1 requires the Mis16-Mis18-Mis19 complex, which is thought to direct recruitment of Scm3-chaperoned Cnp1/histone H4 dimers to DNA. Here, we present the structure of the essential Mis18 partner protein Mis19 and describe its interaction with Mis16, revealing a bipartite-binding site. We provide data on the stoichiometry and overall architecture of the complex and provide detailed insights into the Mis18-Mis19 interface.
© 2019 Korntner-Vetter et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
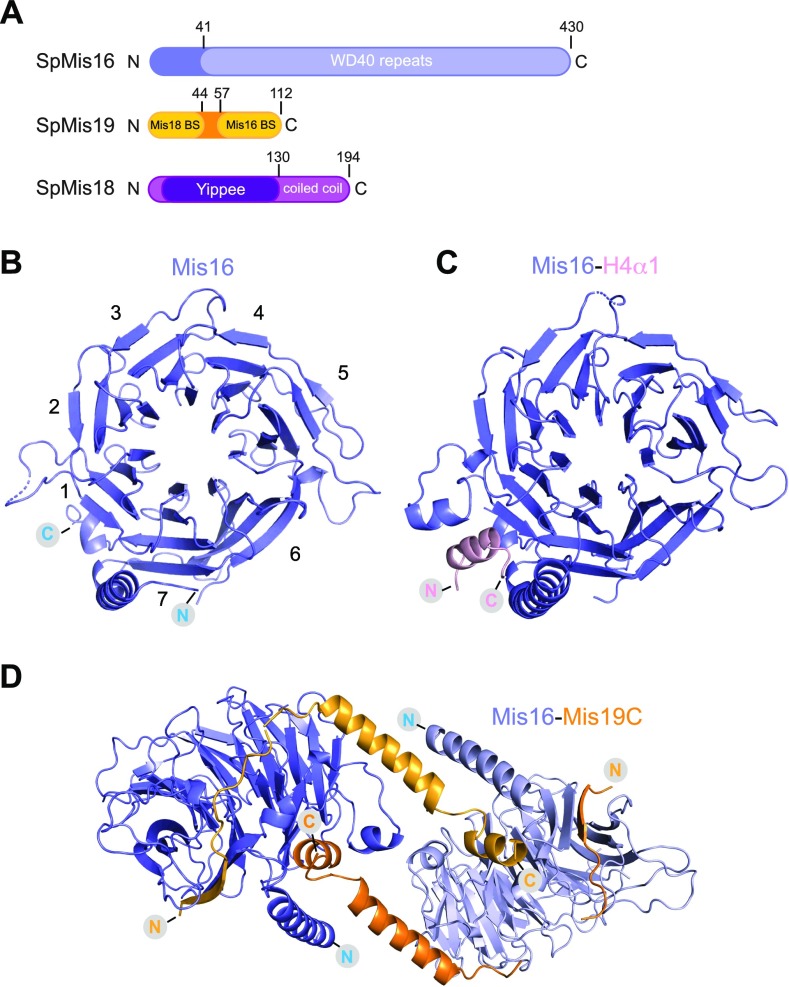
(A) Domain architecture of the S. pombe proteins Mis16, Mis19, and Mis18. Structural domains and putative binding sites are indicated. (B) Crystal structure of the S. pombe Mis16 protein. Propeller blades are numbered. (C) Crystal structure of Mis16 bound to the histone H4 peptide (residues 14–44, ordered residues 28–41). (D) Crystal structure of Mis16 protein bound to the residues 56–111 of Mis19 (orange). The dimer is formed by crystallographic twofold symmetry, and each Mis19 molecule bridges two Mis16 molecules.
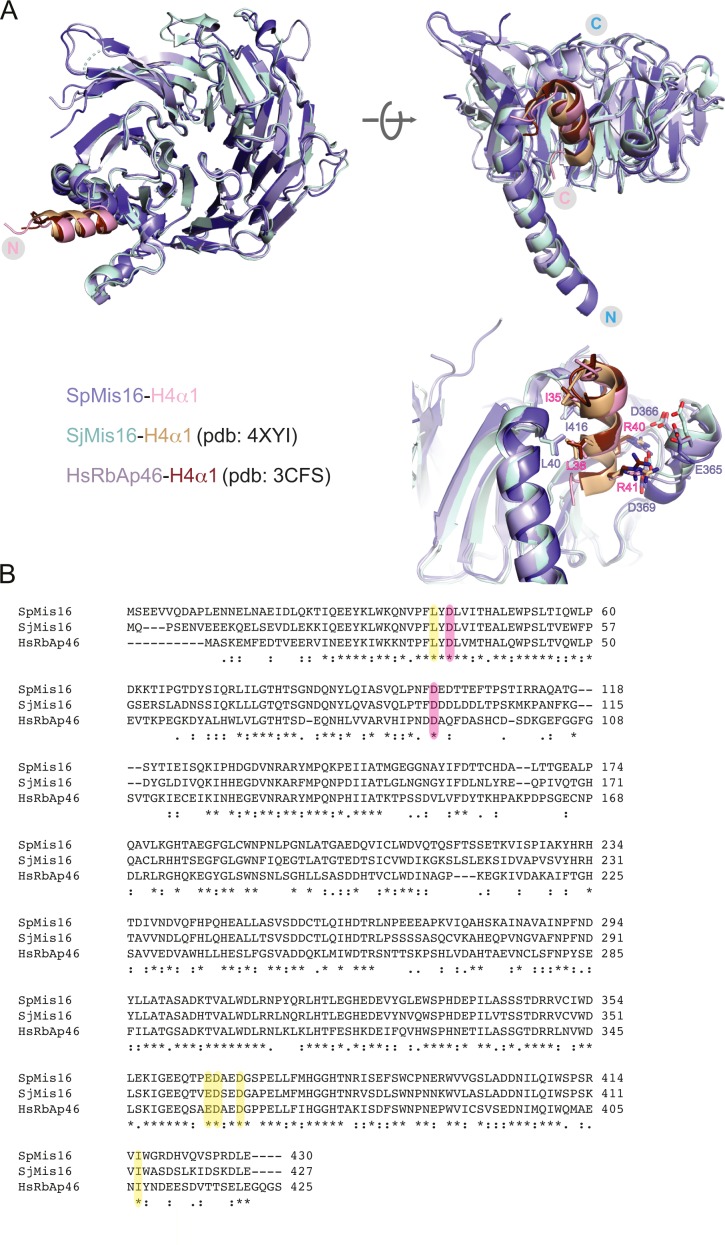
(A) Structural superimposition of the S. pombe Mis16–H4α1 peptide structure with the previously solved crystal structures of S. japonicus Mis16 (PDB ID: 4XYI) and the human Mis16 homologue RbAp46 (PDB ID: 3CFS) bound to H4α1. Inset: A closer view of the Mis16 and H4α1 interaction pocket, showing important residues from Mis16 (blue) and H4 (red). (B) Sequence alignment of Mis16 from S. pombe, S. japonicus and human RbAp46. Identical residues are labelled with an asterisk, similar ones with dots. Residues participating in interactions with histone H4 are highlighted in yellow. Conserved aspartates that bind Mis19 site A are highlighted in pink.

(A) Crystal structure of Mis16–Mis19 highlighting the Mis19-binding sites. Three putative binding sites A, B, and C (lighter to darker orange from N terminus to C terminus) are indicated. (B) Schematic of the various deletion constructs of Mis19 are used for binding studies showing the positions of sites A, B, and C. (C) Constructs containing affinity-tagged Mis16 and the Mis19 deletions shown above were co-expressed and purified by affinity pull-down and size-exclusion chromatography. SDS–PAGE gel of peak fractions is shown. Asterisks indicate impurities. (D) Close-up of the A- and C-binding sites. The locations of the kis1-1 (R65) and eic1-1 (F102) t.s. mutant residues are shown.
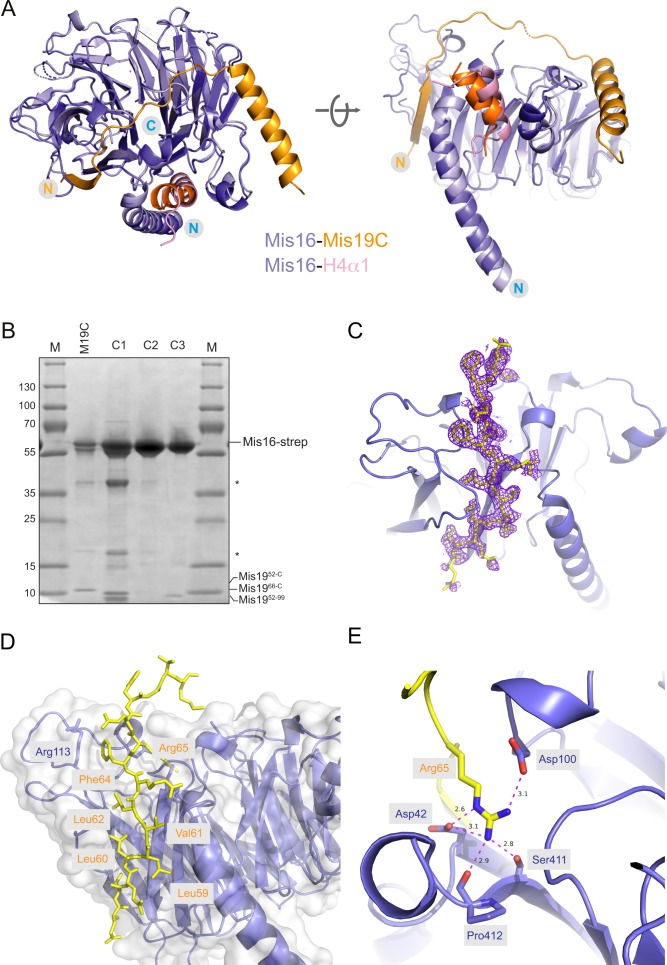
(A) Structural comparison of the S. pombe Mis16–H4α1 and the Mis16–Mis19C complex structure suggest that H4α1 and Mis19 occupy the same binding pocket on Mis16. (B) Replicate pull-down of Mis16 with various Mis19 truncations, confirming results shown in Fig 2C. Nomenclature is the same as in main figure. (C) Electron density (2Fo-Fc) contoured at 1.4σ for the Mis19 A-site (yellow) in the Mis16-binding pocket. (D) Detailed view of the Mis19 A-site showing the run of conserved hydrophobic residues in Mis19 (yellow) immediately preceding the critical Arg65. (E) Close-up view of the interaction between Mis19 Arg65 (yellow) and its binding site in Mis16. Main hydrogen bonding interactions to the Mis16 Asp42/Asp100 side chains and Ser411/Pro412 backbone carbonyl groups are indicated.
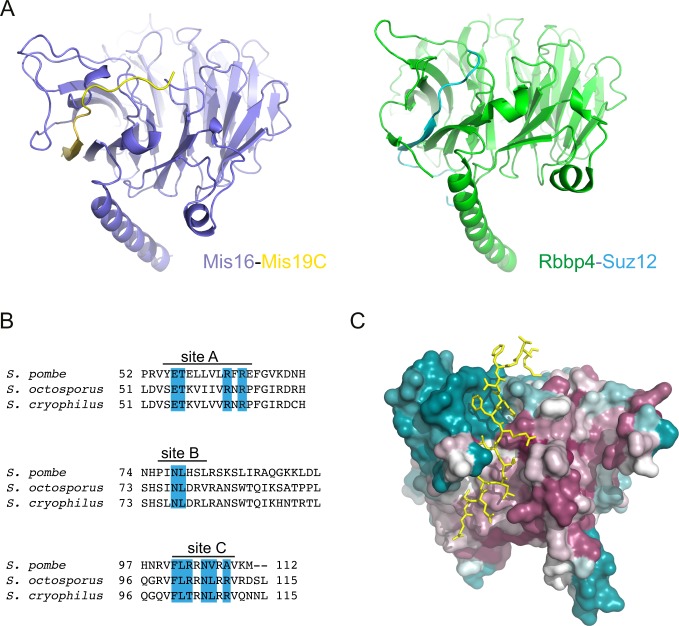
(A) Structural comparison of the Mis16–Mis19 A–binding site, with the interaction between RbAp48 (also known as Rbbp4) and Suz12 observed in the PRC2 complex (PDB ID: 5WAI). (B) Multiple sequence alignment of the C terminus of three Mis19 orthologs. The positions of the putative Mis16-interacting sites are indicated, and identical residues are highlighted in blue. (C) Mis16 surface figure coloured by residue conservation, ranging from high (purple) to low (blue). The bound Mis19 C-site is depicted in yellow.
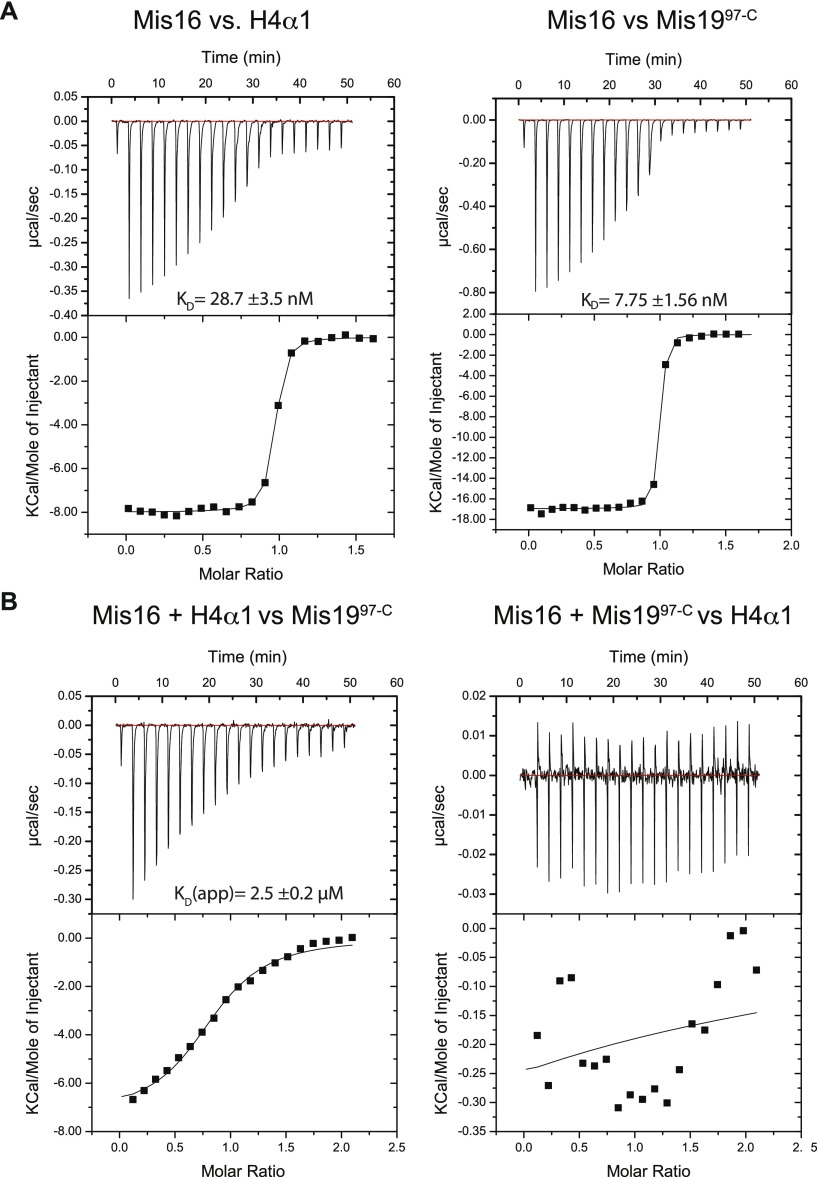
(A) ITC measurements of binding affinity between Mis16 and H4α1 (left) or Mis19 site C (right) peptides. (B) ITC competition experiments. Mis16 was incubated with H4α1 (left) or Mis19 site C (right) peptides, before being challenged with the second substrate.
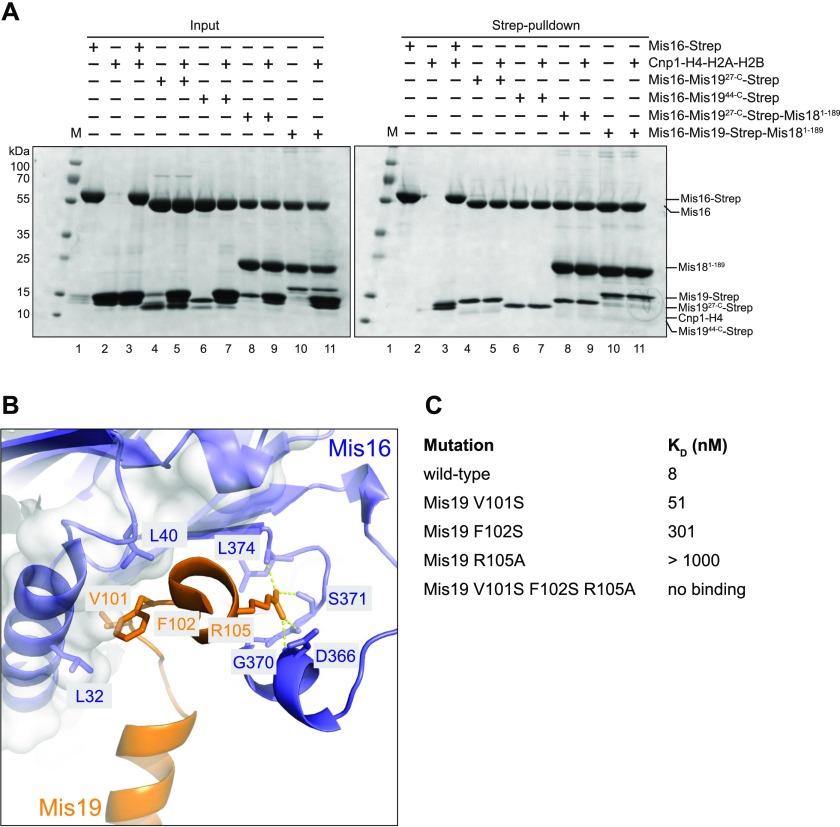
(A) Pull-down assays to test competition between full-length histones Cnp1–H4 and Mis16–19 or Mis16–18–19 complexes. Left panel shows input SDS–PAGE gel of purified proteins and right panel shows results of pull-down assay. (B) Location of key residues in the Mis16–H4α1 interaction. (C) Binding constants of point and triple mutants of the Mis19 C-terminal peptide (residues 97–112) against Mis16, as determined by ITC.
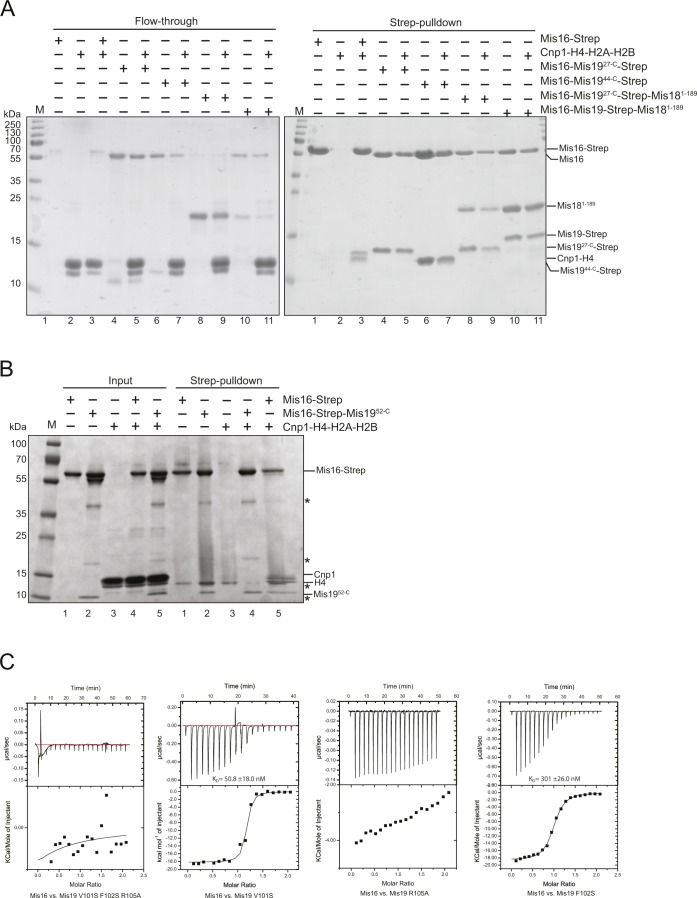
(A) Repeat pull-down of the Mis16 competition experiments shown in Fig 4A. Left panel shows unbound input; right panel shows pull-downs. (B) Pull-down of histones Cnp1–H4 by Strep-tagged Mis16 in the absence and presence of untagged Mis1952-C. Inputs are shown on the left and pull-downs on the right. In the presence of Mis1952-C, no histones can be transferred onto Strep-Mis16. Asterisks indicate impurities. (C) Raw ITC data and fitted binding isotherms for the Mis16–Mis19 mutant–binding studies.
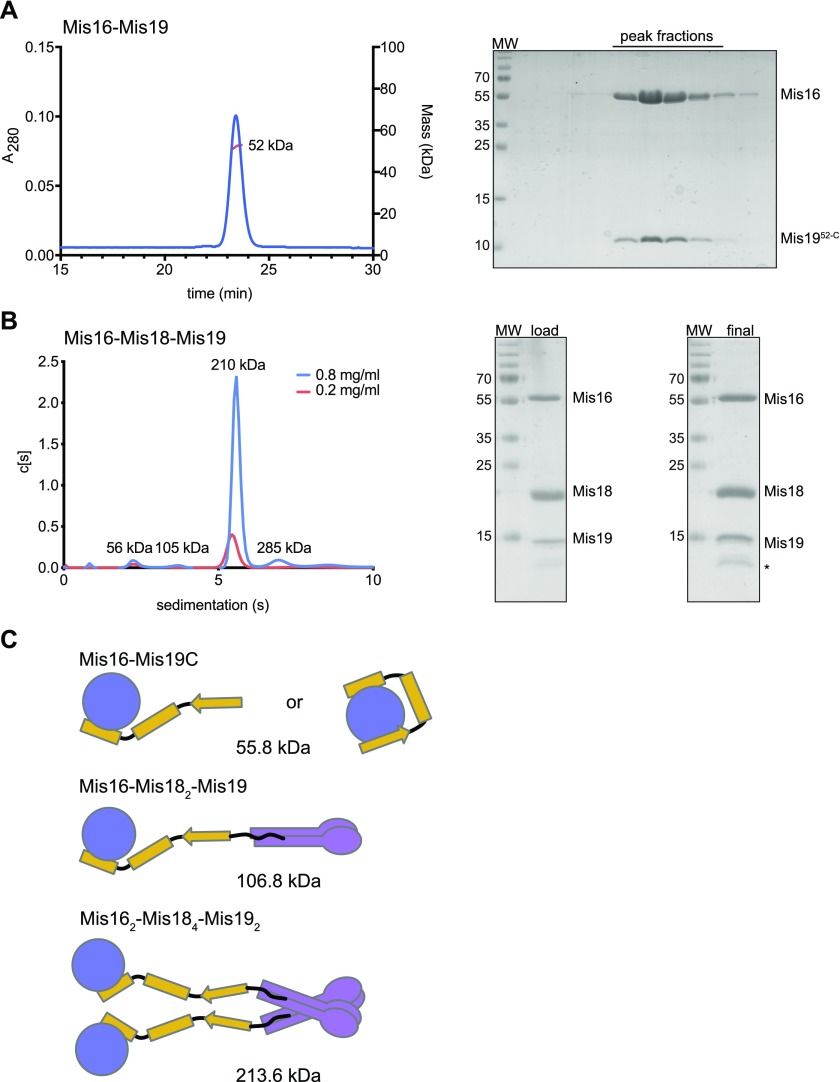
(A) MALLS measurement of mass of the Mis16–Mis1952-C complex. Right panel shows SDS–PAGE gel of elution fractions. (B) AUC analysis of the Mis16–Mis18–Mis19 complex at two concentrations (indicated). Right panel shows SDS–PAGE gels of input and recovered protein. (C) Schematic showing possible subunit arrangements that account for the observed solution masses. Different configurations are possible for the Mis16–Mis19 interaction depending on whether one or two binding sites are occupied, as indicated in the figure. Similar interactions could also occur in the larger complex but are omitted for clarity. Calculated masses are indicated for the untagged proteins.
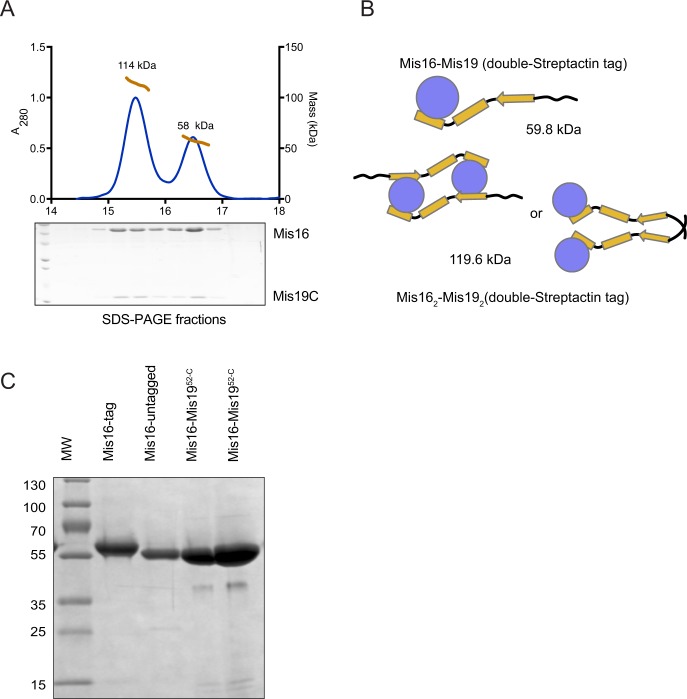
(A) MALLS analysis of double-Strep-Tactin–tagged Mis16–Mis1952-C complex (tag on Mis16). The trace shows the presence of two species, corresponding to a heterodimer and dimer-of-heterodimers. SDS–PAGE analysis shows the presence of both proteins in each peak. (B) Schematic diagram of the possible oligomerisation and calculated masses of the complexes in the MALLS peaks. The dimeric Mis16–Mis19 may also bind as shown in Fig 5C. (C) SDS–PAGE gel of the Mis16–Mis19C complex used for crystallization compared with tagged and untagged Mis16 alone. The tag is completely removed from the crystallised sample.
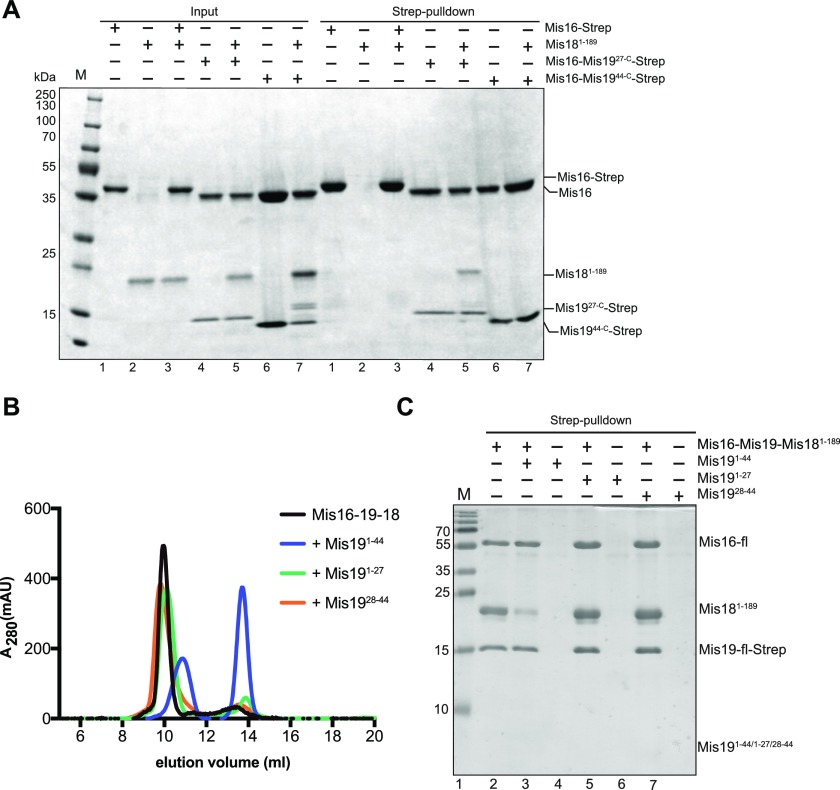
(A) Mis16–Mis18–Mis19 interactions assessed by pull-down. Input proteins are shown on the left. Binding of Mis16–Mis19 and Mis19 truncations to Mis18 are assessed by pull-down of Mis19 (right). (B) Mis16–19–18 binding assessed by size-exclusion and competition assays. Preformed Mis16–18–19 complex was challenged with peptides derived from the N terminus of Mis19 (indicated). Competitive interactions result in the dissociation of the complex into separate Mis16–19 and Mis18 peaks. (C) Pull-down assay of Mis16–Mis19–Strep–Mis18 complex in the absence (lane 1) and the presence of Mis191–44 (lane 2), Mis191–27 (lane 4), and Mis1928–44 (lane 6) peptides.
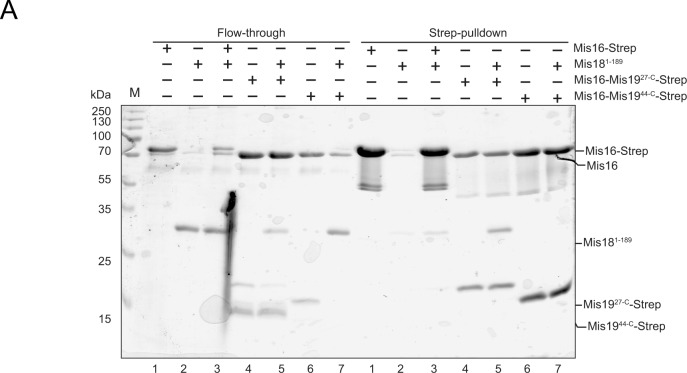
(A) Interactions of Mis16–19 with Mis18 (repeat of pull-down shown in Fig 6A). Left panel shows unbound input; right panel shows pull-downs.
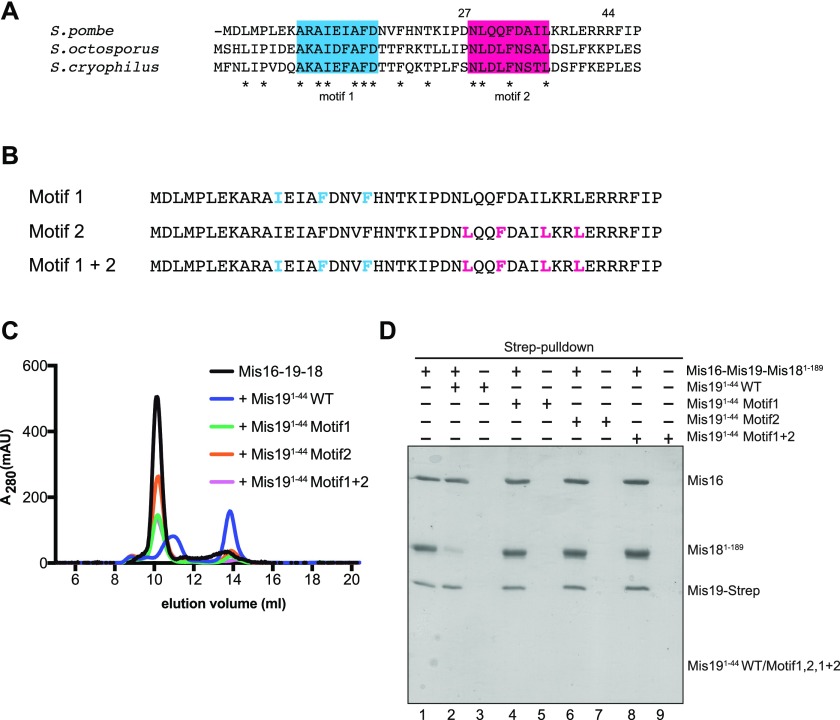
(A) Multiple sequence alignment of the N terminus of Mis19 orthologs. Two putative conserved regions were identified (blue and red). Asterisks indicate totally conserved residues. (B) Peptides used for competition assays containing conserved hydrophobic residues in the Mis19 N terminus. Residues in motif 1 (blue) or motif 2 (pink) were selected for alanine mutagenesis and competition binding analysis. (C) Repeat of competition assays in (6B) using peptides with alanine mutations in motif 1, motif 2 or both motifs. (D) Repeat of pull-down assays in (6C) using peptides with alanine mutations in motif 1, motif 2, or both motifs.
References
-
- Adams PD, Afonine PV, Bunkóczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung L-W, Kapral GJ, Grosse-Kunstleve RW, et al. (2010) PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr 66: 213–221. 10.1107/s0907444909052925 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases