

Site-specific phosphorylation of Fbxw7 by Cdk5/p25 and its resulting decreased stability are linked to glutamate-induced excitotoxicity
- PMID: 31371703
- PMCID: PMC6675790
- DOI: 10.1038/s41419-019-1818-4
Site-specific phosphorylation of Fbxw7 by Cdk5/p25 and its resulting decreased stability are linked to glutamate-induced excitotoxicity
Abstract
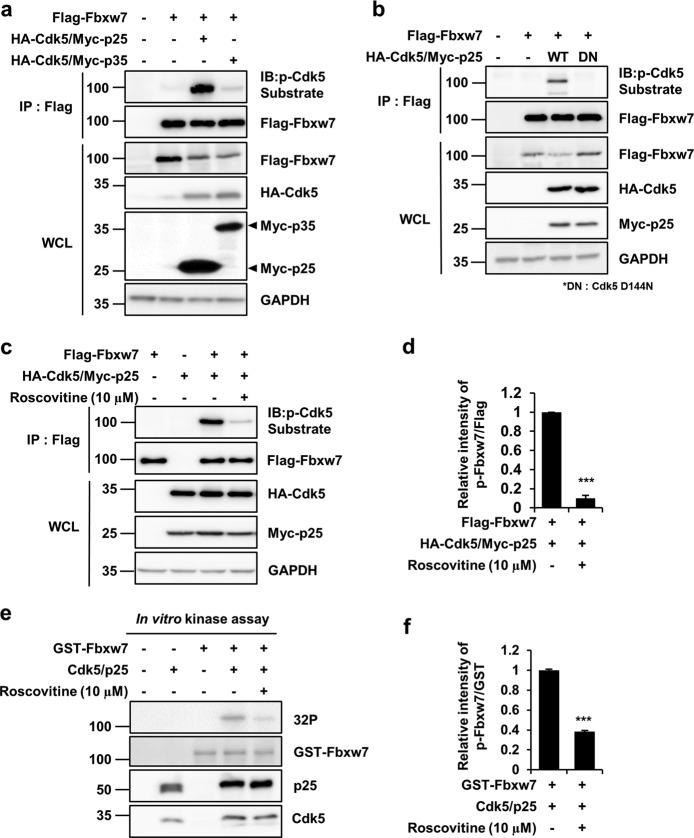
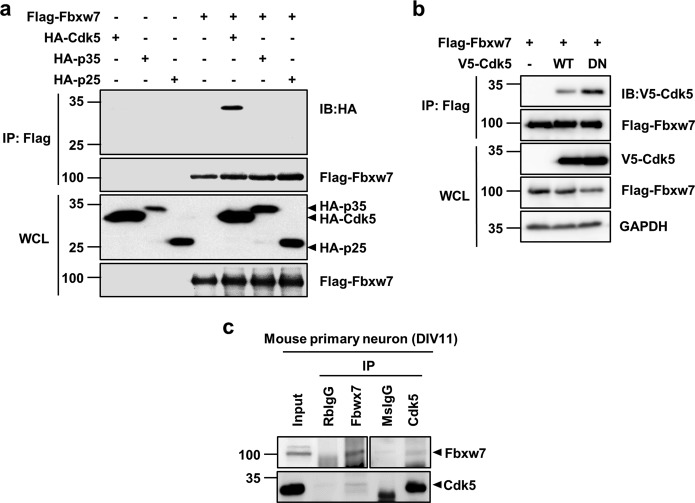
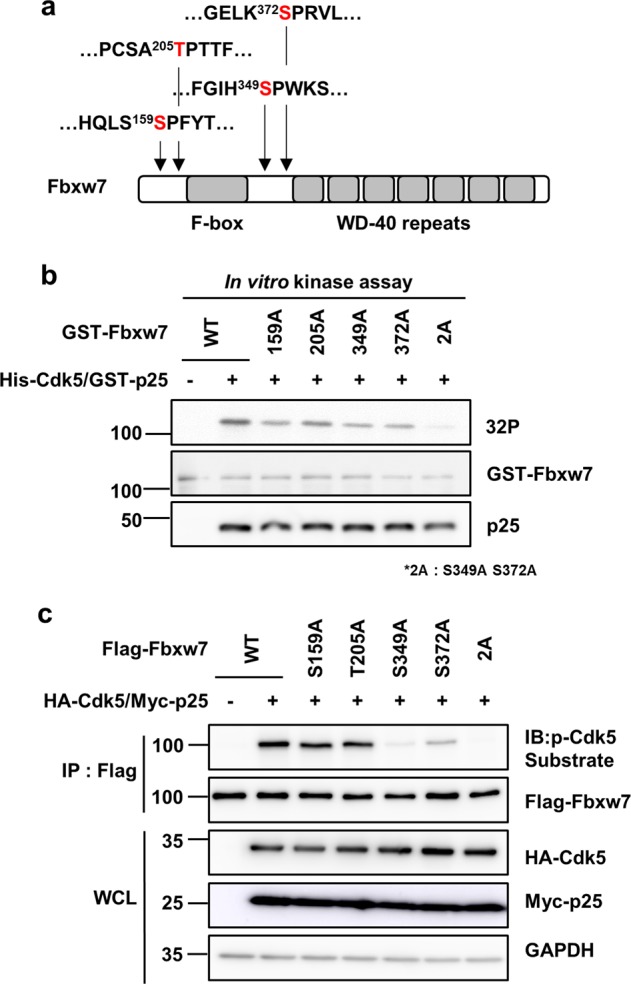
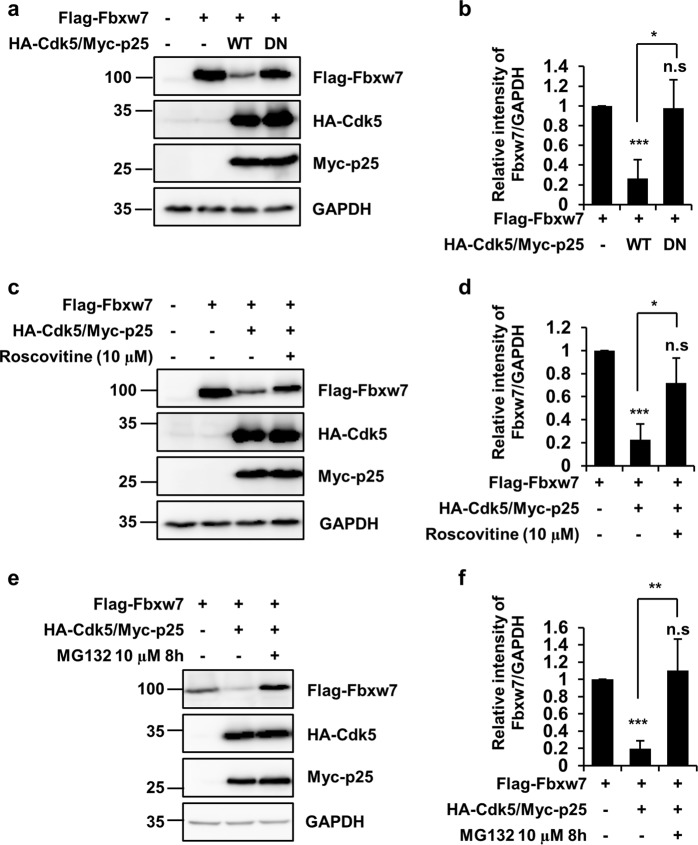
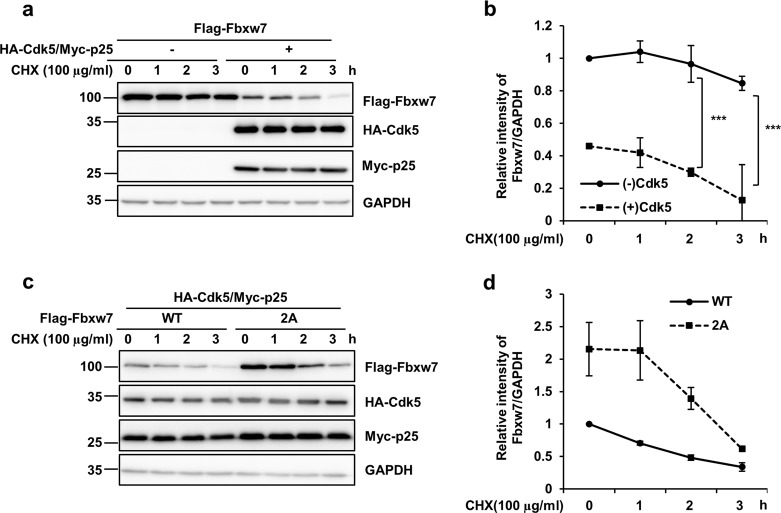
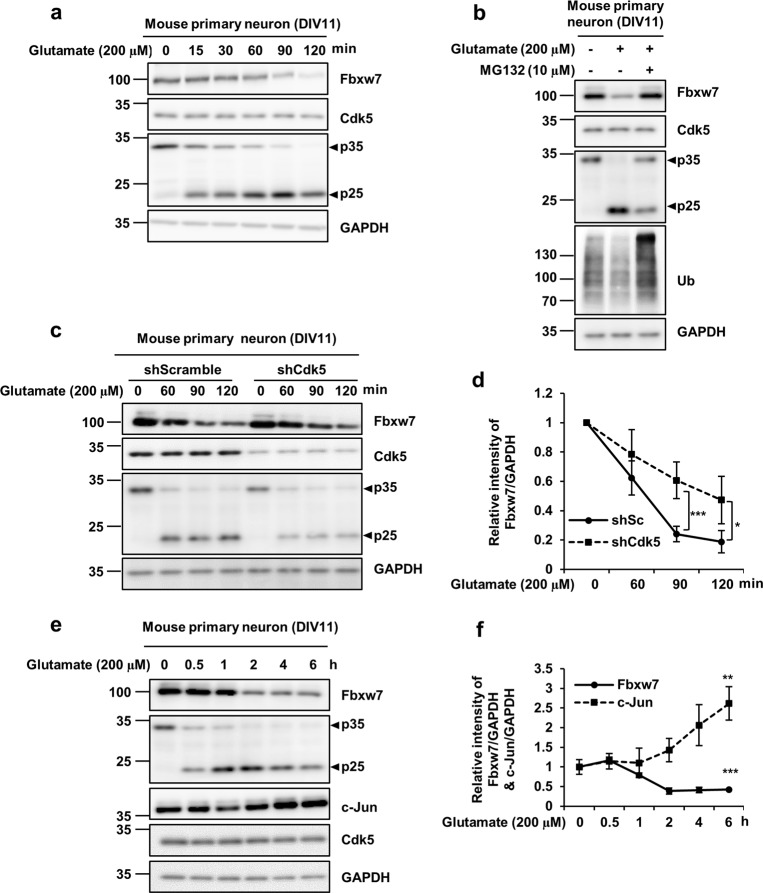
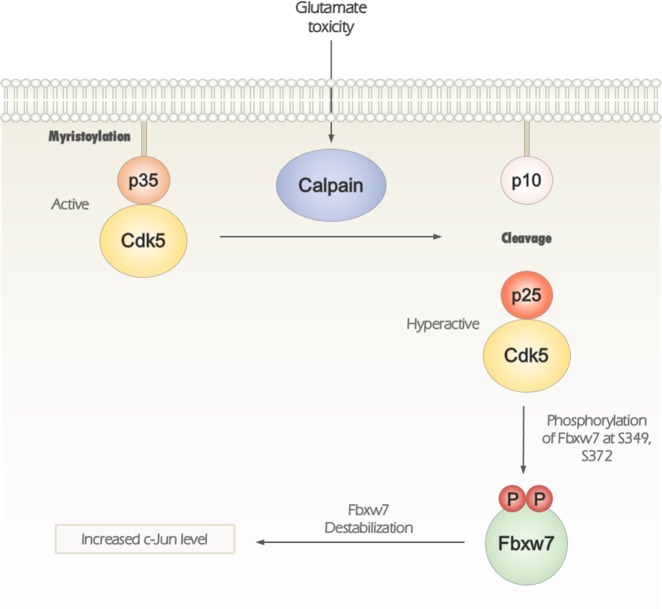
Cyclin-dependent kinase 5 (Cdk5) is a serine/threonine protein kinase that regulates brain development and neurodegeneration. Cdk5 is activated by p25 that is generated from calpain-dependent cleavage of p35. The generation of p25 is responsible for the aberrant hyper-activation of Cdk5, which causes neurodegeneration. Using in vitro assays, we discovered that F-box/WD repeat-containing protein 7 (Fbxw7) is a new substrate of Cdk5. Additionally, Cdk5-dependent phosphorylation of Fbxw7 was detected in the presence of p25, and two amino acid residues (S349 and S372) were determined to be major phosphorylation sites. This phosphorylation was eventually linked to decreased stability of Fbxw7. Using a culture model of cortical neurons challenged with glutamate, we confirmed that decreased stability of Fbxw7 was indeed Cdk5-dependent. Furthermore, diminished levels of Fbxw7 led to increased levels of transcription factor AP-1 (c-Jun), a known substrate of Fbxw7. Given that previous reports demonstrate that c-Jun plays a role in accelerating neuronal apoptosis in these pathological models, our data support the concepts of a molecular cascade in which Cdk5-mediated phosphorylation of Fbxw7 negatively regulates Fbxw7 expression, thereby contributing to neuronal cell death following glutamate-mediated excitotoxicity.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
