

Engineering Orthogonal Polypeptide GalNAc-Transferase and UDP-Sugar Pairs
- PMID: 31373799
- PMCID: PMC6813768
- DOI: 10.1021/jacs.9b04695
Engineering Orthogonal Polypeptide GalNAc-Transferase and UDP-Sugar Pairs
Abstract
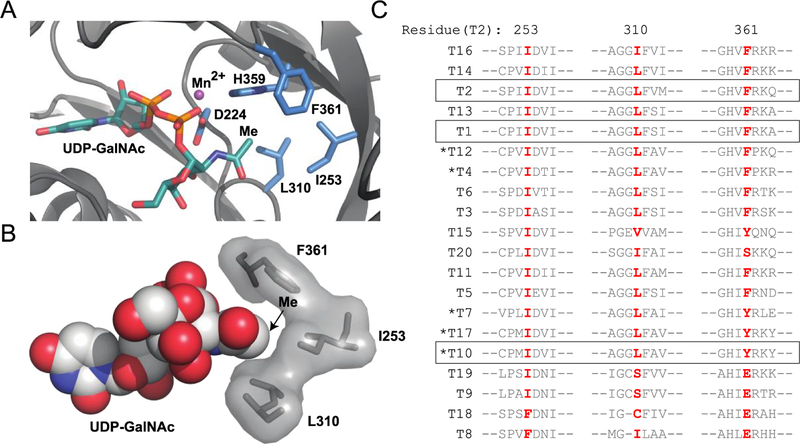
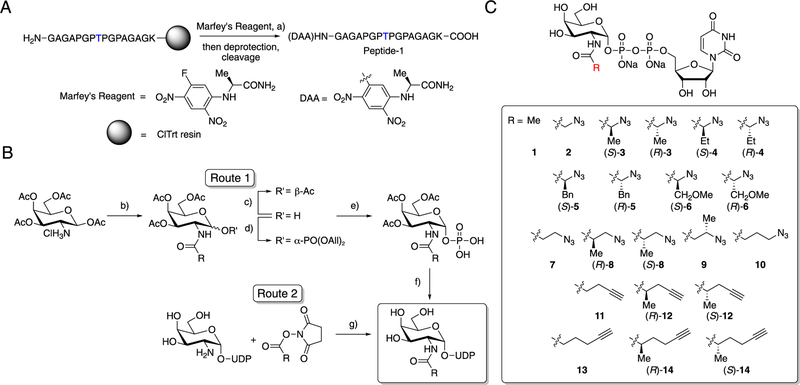
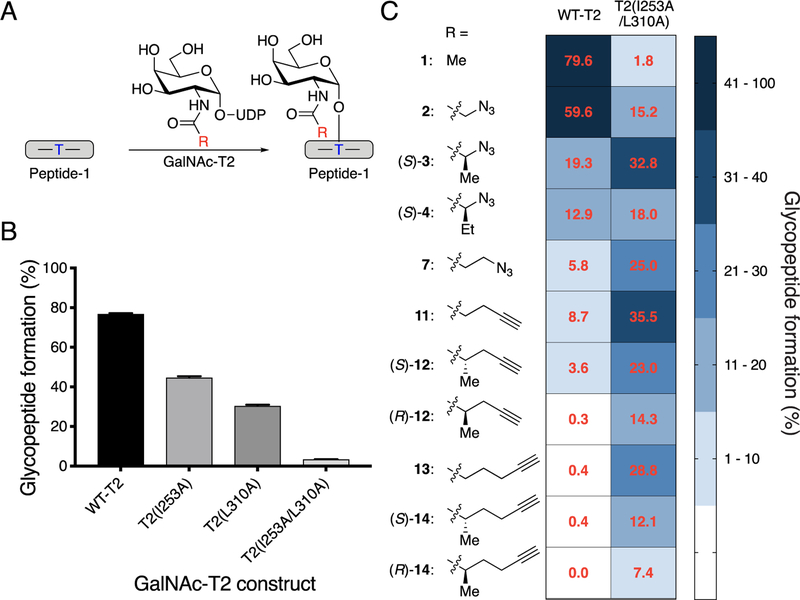
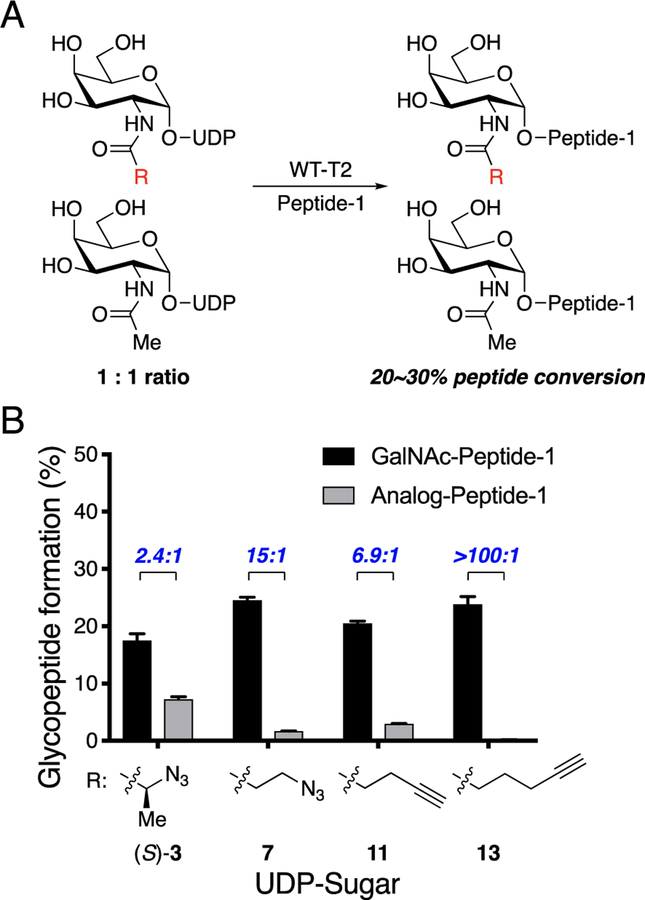
O-Linked α-N-acetylgalactosamine (O-GalNAc) glycans constitute a major part of the human glycome. They are difficult to study because of the complex interplay of 20 distinct glycosyltransferase isoenzymes that initiate this form of glycosylation, the polypeptide N-acetylgalactosaminyltransferases (GalNAc-Ts). Despite proven disease relevance, correlating the activity of individual GalNAc-Ts with biological function remains challenging due to a lack of tools to probe their substrate specificity in a complex biological environment. Here, we develop a "bump-hole" chemical reporter system for studying GalNAc-T activity in vitro. Individual GalNAc-Ts were rationally engineered to contain an enlarged active site (hole) and probed with a newly synthesized collection of 20 (bumped) uridine diphosphate N-acetylgalactosamine (UDP-GalNAc) analogs to identify enzyme-substrate pairs that retain peptide specificities but are otherwise completely orthogonal to native enzyme-substrate pairs. The approach was applicable to multiple GalNAc-T isoenzymes, including GalNAc-T1 and -T2 that prefer nonglycosylated peptide substrates and GalNAcT-10 that prefers a preglycosylated peptide substrate. A detailed investigation of enzyme kinetics and specificities revealed the robustness of the approach to faithfully report on GalNAc-T activity and paves the way for studying substrate specificities in living systems.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
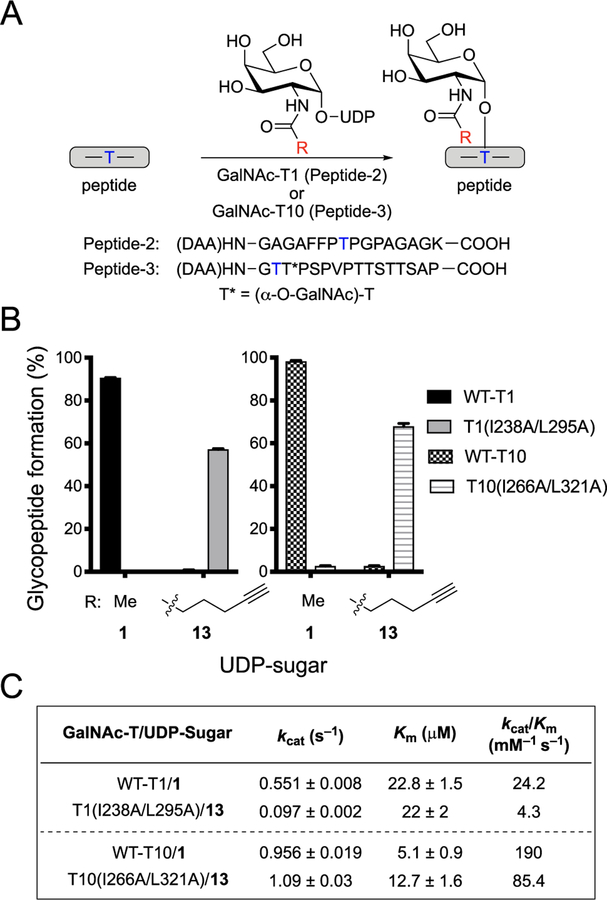
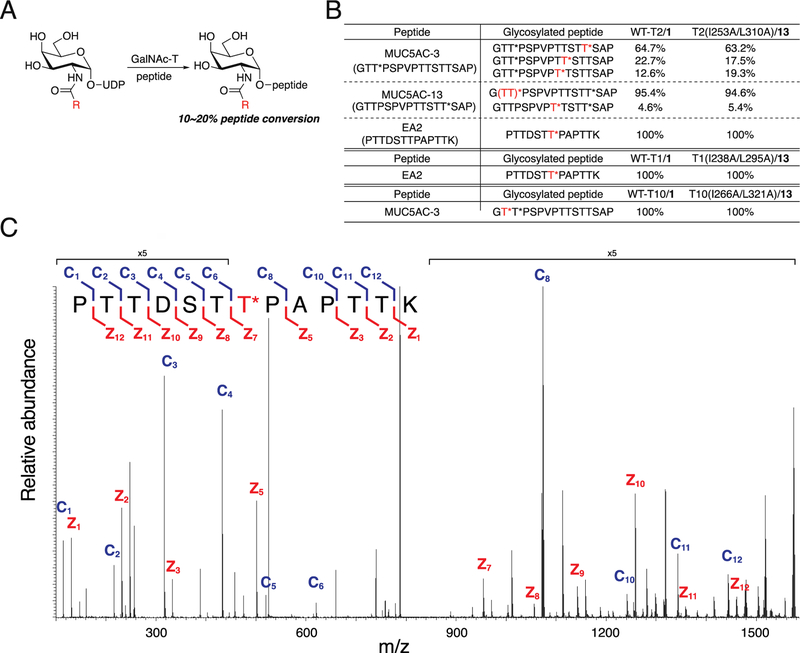
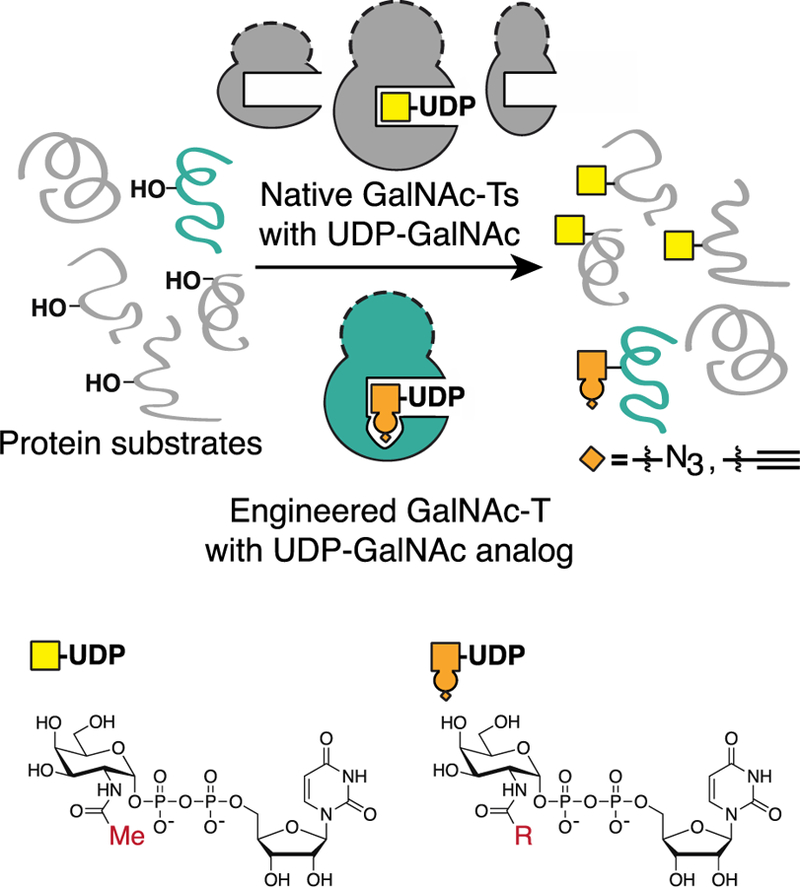
References
-
- Xu Z; Weiss A Negative Regulation of CD45 by Differential Homodimerization of the Alternatively Spliced Isoforms. Nat. Immunol 2002, 3 (8), 764–771. - PubMed
-
- Radhakrishnan P; Dabelsteen S; Madsen FB; Francavilla C; Kopp KL; Steentoft C; Vakhrushev SY; Olsen JV; Hansen L; Bennett EP; Woetmann A; Yin G; Chen L; Song H; Bak M; Hlady RA; Peters SL; Opavsky R; Thode C; Qvortrup K; Schjoldager KT-BG; Clausen H; Hollingsworth MA; Wandall HH Immature Truncated O-Glycophenotype of Cancer Directly Induces Oncogenic Features. Proc. Natl. Acad. Sci. U. S. A 2014, 111 (39), E4066–E4075. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
