

Evolution of fast root gravitropism in seed plants
- PMID: 31375675
- PMCID: PMC6677796
- DOI: 10.1038/s41467-019-11471-8
Evolution of fast root gravitropism in seed plants
Abstract
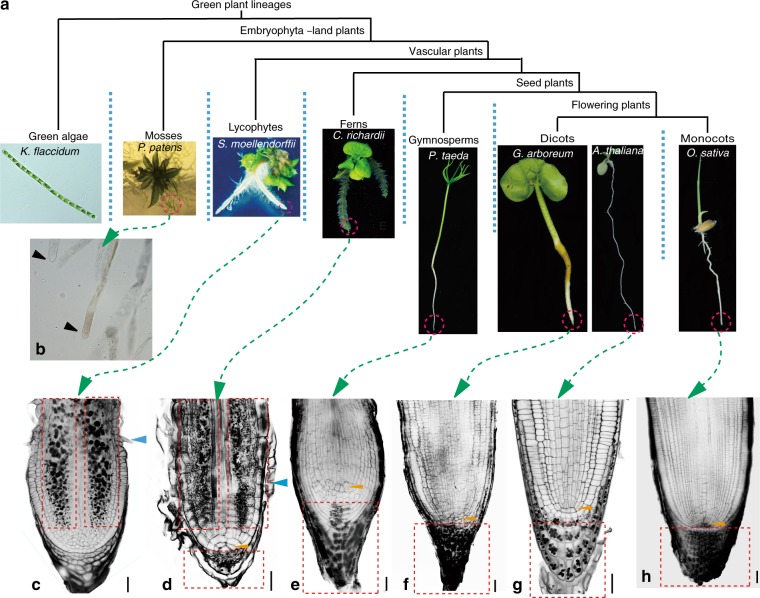
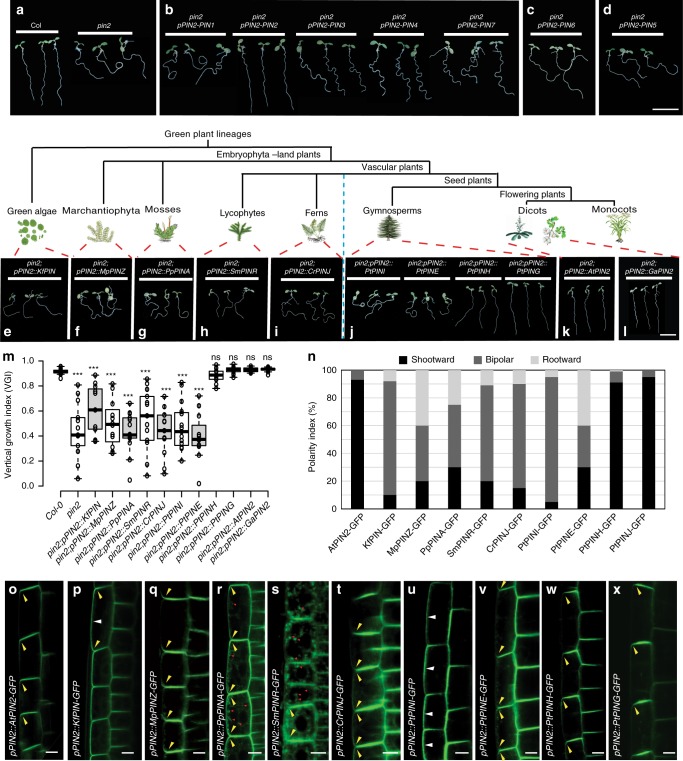
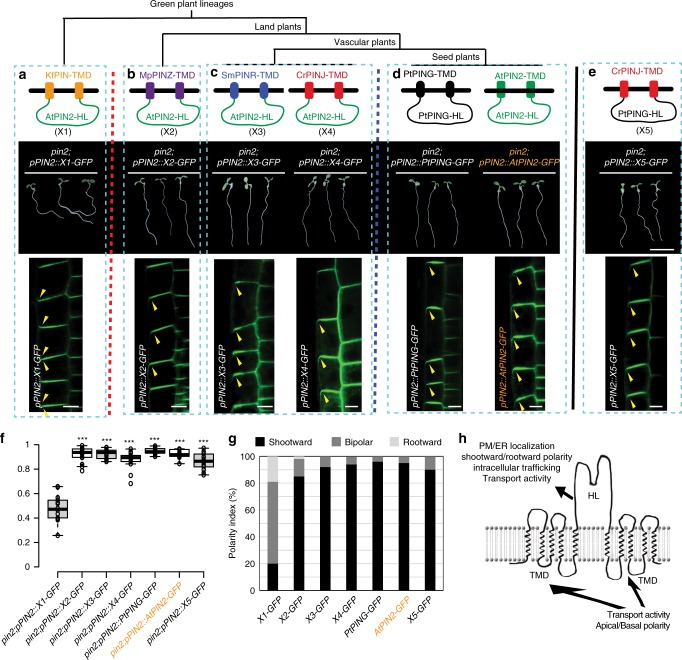
An important adaptation during colonization of land by plants is gravitropic growth of roots, which enabled roots to reach water and nutrients, and firmly anchor plants in the ground. Here we provide insights into the evolution of an efficient root gravitropic mechanism in the seed plants. Architectural innovation, with gravity perception constrained in the root tips along with a shootward transport route for the phytohormone auxin, appeared only upon the emergence of seed plants. Interspecies complementation and protein domain swapping revealed functional innovations within the PIN family of auxin transporters leading to the evolution of gravitropism-specific PINs. The unique apical/shootward subcellular localization of PIN proteins is the major evolutionary innovation that connected the anatomically separated sites of gravity perception and growth response via the mobile auxin signal. We conclude that the crucial anatomical and functional components emerged hand-in-hand to facilitate the evolution of fast gravitropic response, which is one of the major adaptations of seed plants to dry land.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Langdale, J. A. & Harrison, C. J. in Fusco AMAG. Evolving Pathways: Key Themes in Evolutionary Developmental Biology p. 299–315 (Cambridge Univ. Press, Cambridge, MA, 2008).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
