

X-ray Crystallographic Structure and Oligomerization of Gloeobacter Rhodopsin
- PMID: 31375689
- PMCID: PMC6677831
- DOI: 10.1038/s41598-019-47445-5
X-ray Crystallographic Structure and Oligomerization of Gloeobacter Rhodopsin
Abstract
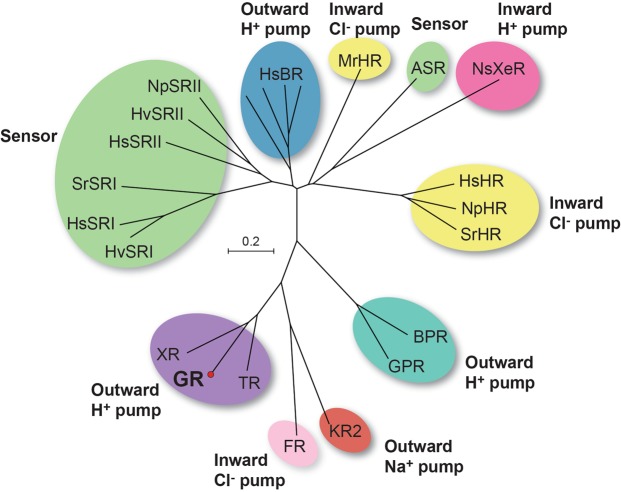
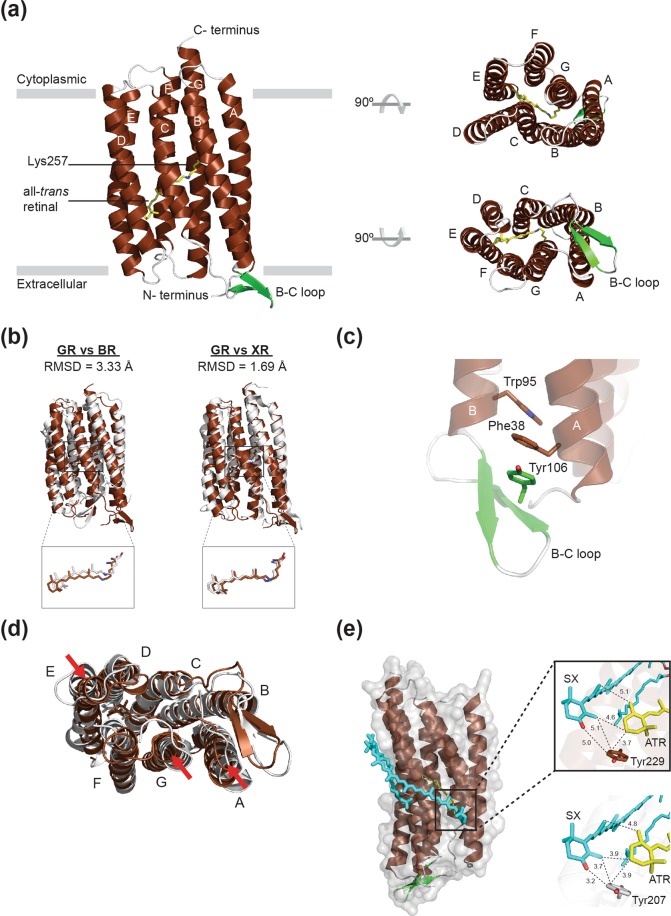
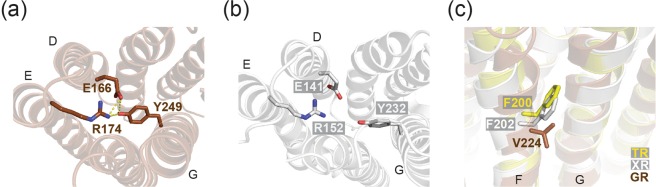
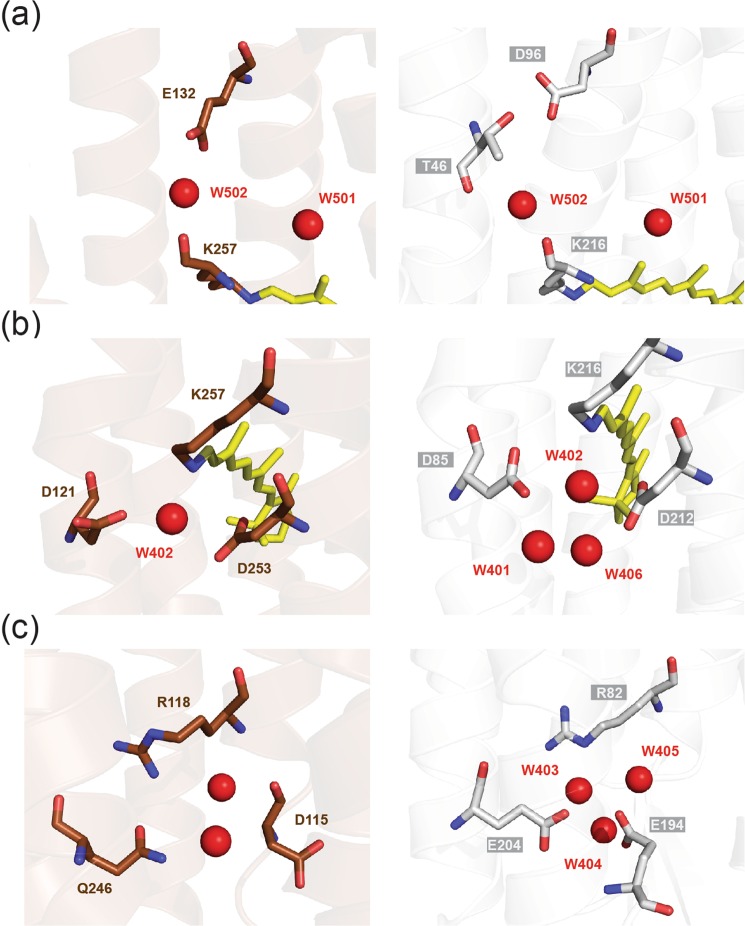
Gloeobacter rhodopsin (GR) is a cyanobacterial proton pump which can be potentially applied to optogenetics. We solved the crystal structure of GR and found that it has overall similarity to the homologous proton pump from Salinibacter ruber, xanthorhodopsin (XR). We identified distinct structural characteristics of GR's hydrogen bonding network in the transmembrane domain as well as the displacement of extracellular sides of the transmembrane helices relative to those of XR. Employing Raman spectroscopy and flash-photolysis, we found that GR in the crystals exists in a state which displays retinal conformation and photochemical cycle similar to the functional form observed in lipids. Based on the crystal structure of GR, we selected a site for spin labeling to determine GR's oligomerization state using double electron-electron resonance (DEER) spectroscopy and demonstrated the pH-dependent pentamer formation of GR. Determination of the structure of GR as well as its pentamerizing propensity enabled us to reveal the role of structural motifs (extended helices, 3-omega motif and flipped B-C loop) commonly found among light-driven bacterial pumps in oligomer formation. Here we propose a new concept to classify these pumps based on the relationship between their oligomerization propensities and these structural determinants.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Spudich, J. L., Yang, C. S., Jung, K. H. & Spudich, E. N. Retinylidene Proteins: Structures and Functions from Archaea to Humans. Annual review of cell and developmental biology16 (2000). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
