

Fatty Acid Metabolites Combine with Reduced β Oxidation to Activate Th17 Inflammation in Human Type 2 Diabetes
- PMID: 31378464
- PMCID: PMC8506657
- DOI: 10.1016/j.cmet.2019.07.004
Fatty Acid Metabolites Combine with Reduced β Oxidation to Activate Th17 Inflammation in Human Type 2 Diabetes
Abstract
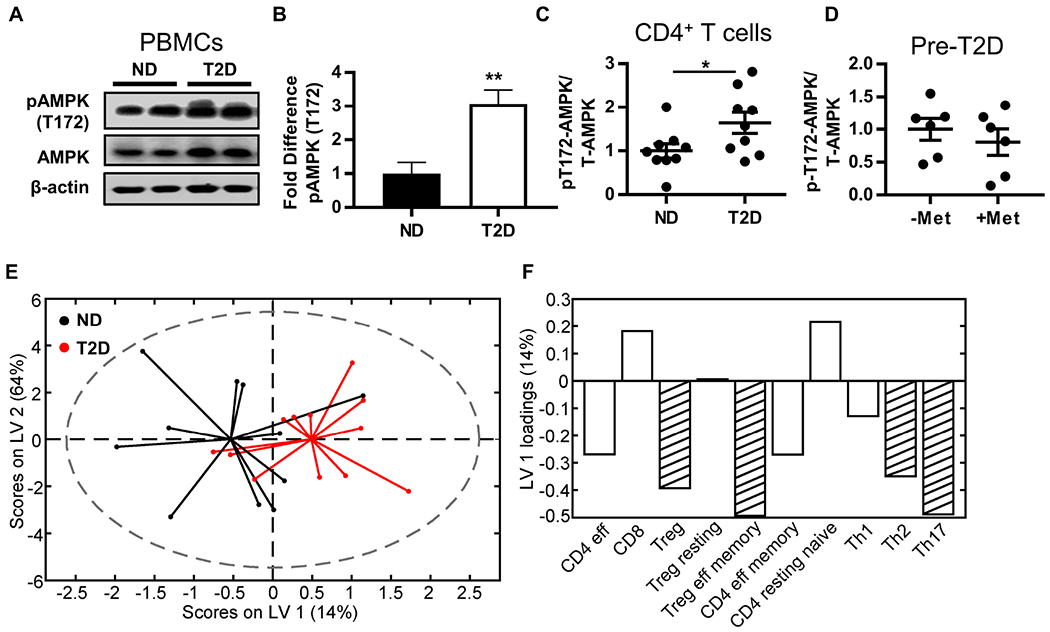
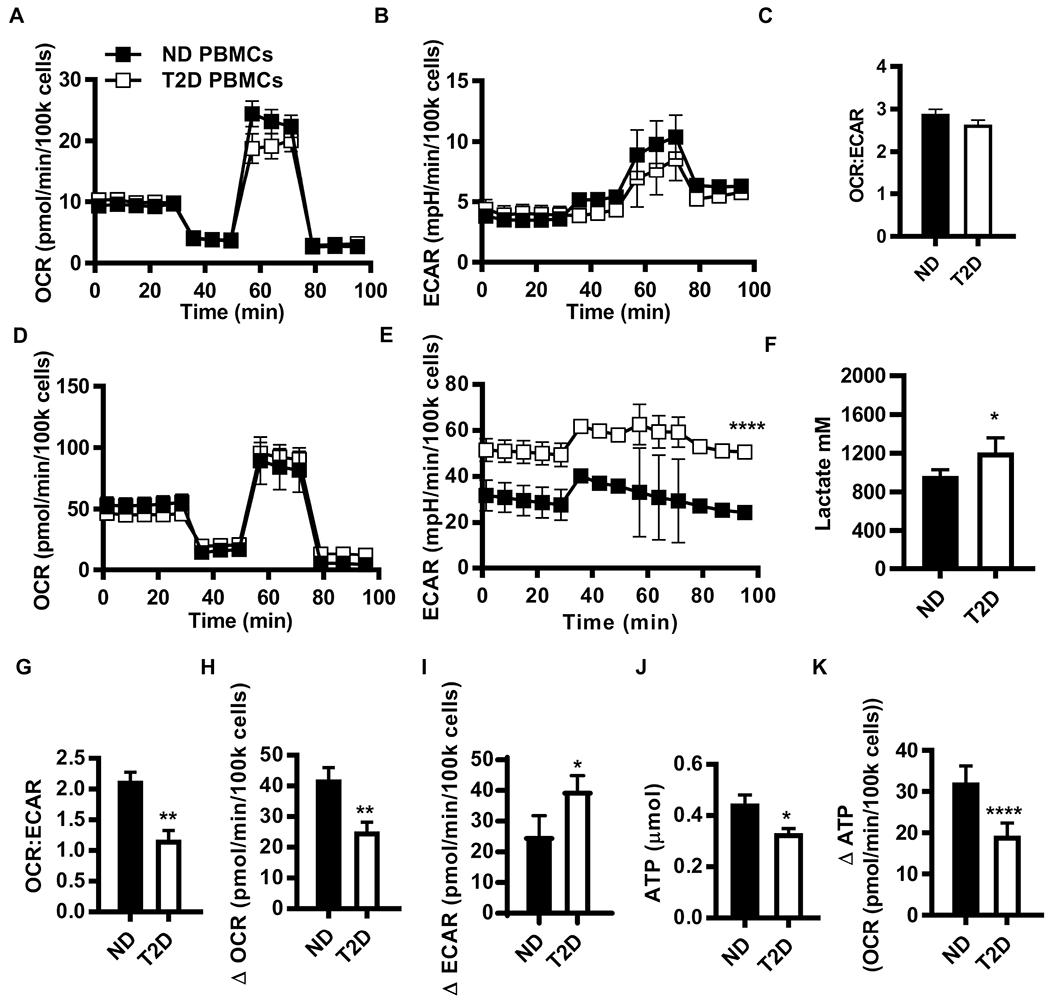
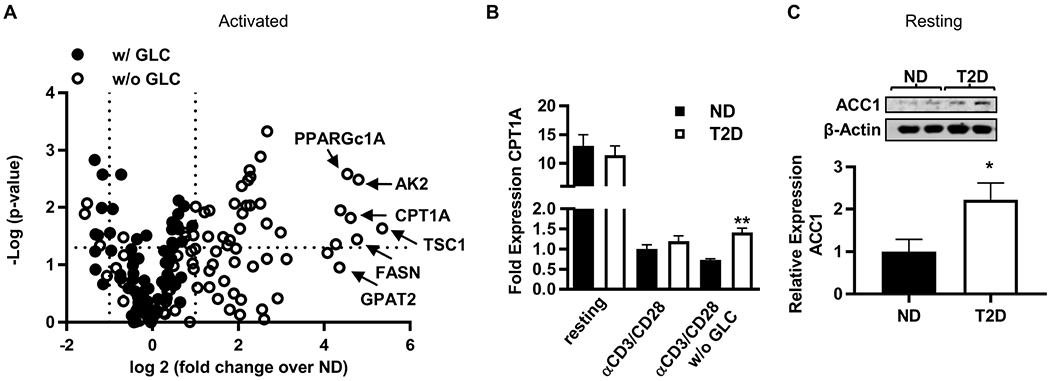
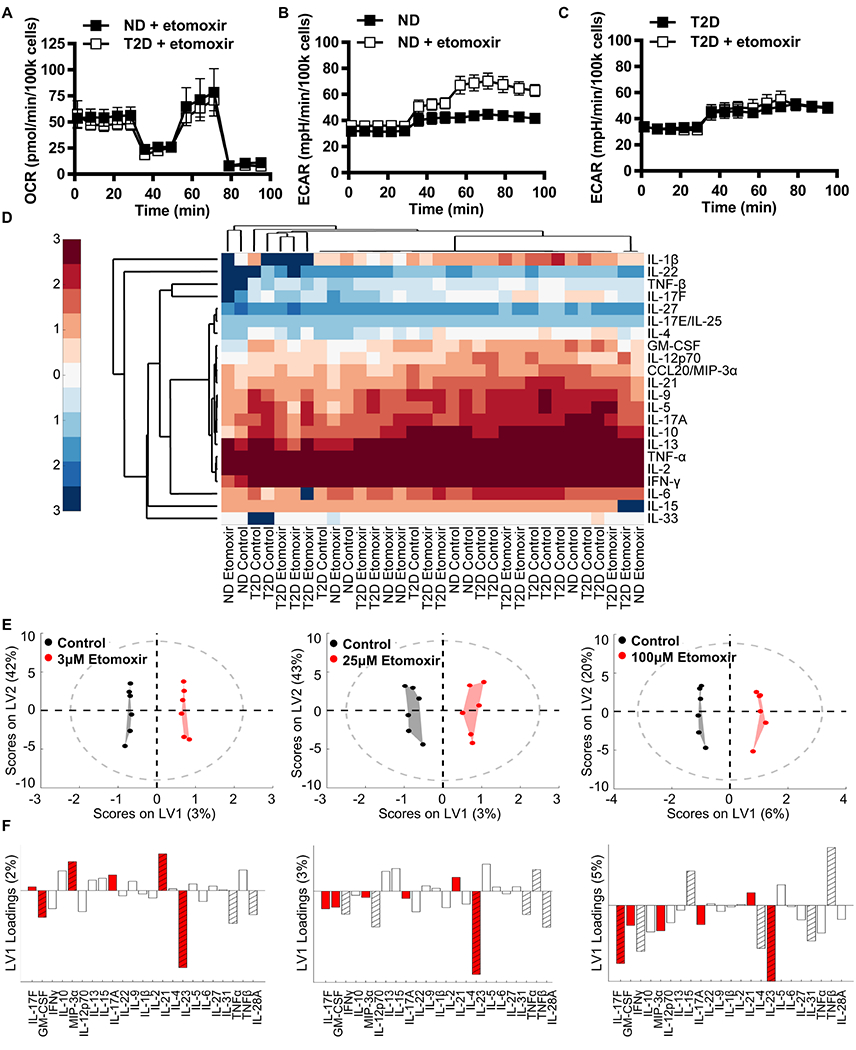
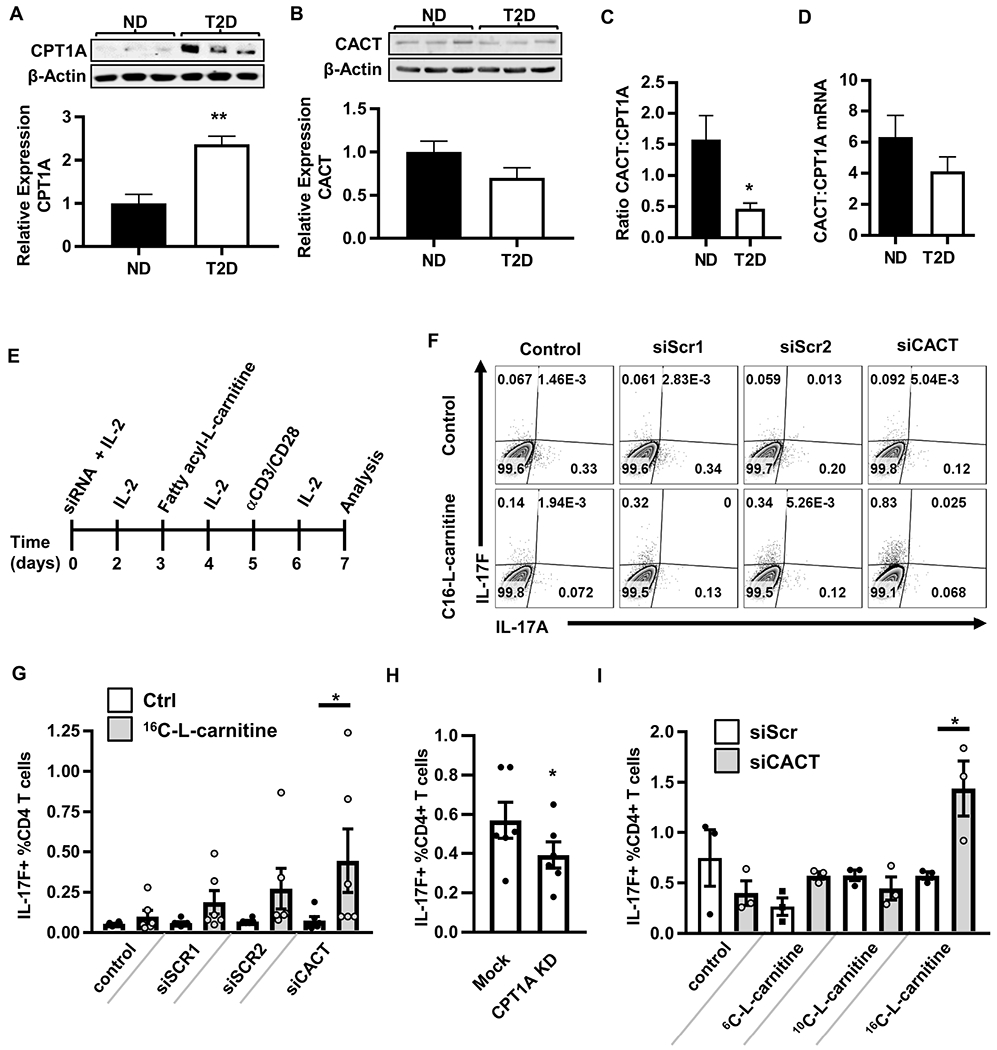
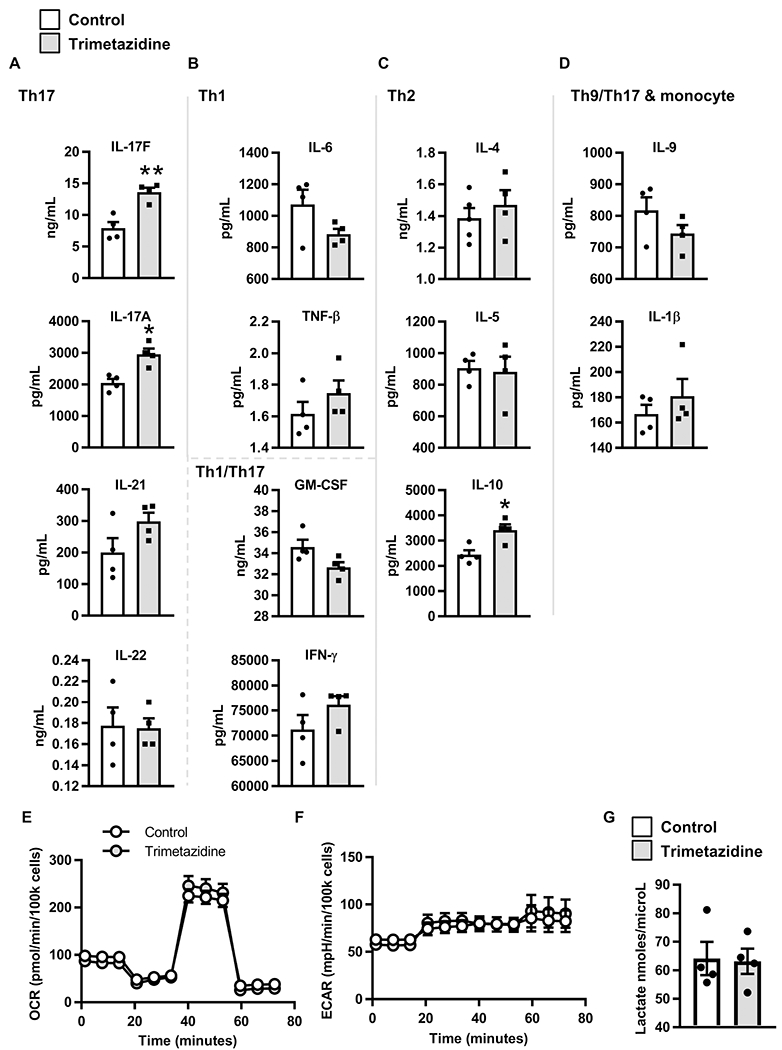
Mechanisms that regulate metabolites and downstream energy generation are key determinants of T cell cytokine production, but the processes underlying the Th17 profile that predicts the metabolic status of people with obesity are untested. Th17 function requires fatty acid uptake, and our new data show that blockade of CPT1A inhibits Th17-associated cytokine production by cells from people with type 2 diabetes (T2D). A low CACT:CPT1A ratio in immune cells from T2D subjects indicates altered mitochondrial function and coincides with the preference of these cells to generate ATP through glycolysis rather than fatty acid oxidation. However, glycolysis was not critical for Th17 cytokines. Instead, β oxidation blockade or CACT knockdown in T cells from lean subjects to mimic characteristics of T2D causes cells to utilize 16C-fatty acylcarnitine to support Th17 cytokines. These data show long-chain acylcarnitine combines with compromised β oxidation to promote disease-predictive inflammation in human T2D.
Keywords: fatty acid oxidation; glycolysis; immunometabolism; metaflammation.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Competing Interests: The authors have no competing interests relevant to the work.
Figures

Comment in
-
Glucose isn't always to blame.Nat Rev Endocrinol. 2019 Oct;15(10):564. doi: 10.1038/s41574-019-0252-0. Nat Rev Endocrinol. 2019. PMID: 31417193 No abstract available.
References
-
- Agilent Technologies, I. (2017). Agilent Seahorse XF Mito Fuel Flex Test Kit User Manual. (Wilmington, DE: ).
-
- Blagih J, Coulombe F, Vincent EE, Dupuy F, Galicia-Vazquez G, Yurchenko E, Raissi TC, van der Windt GJ, Viollet B, Pearce EL, et al. (2015). The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity 42, 41–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
