

CRISPR/Cas9 Delivery Mediated with Hydroxyl-Rich Nanosystems for Gene Editing in Aorta
- PMID: 31380173
- PMCID: PMC6662060
- DOI: 10.1002/advs.201900386
CRISPR/Cas9 Delivery Mediated with Hydroxyl-Rich Nanosystems for Gene Editing in Aorta
Abstract
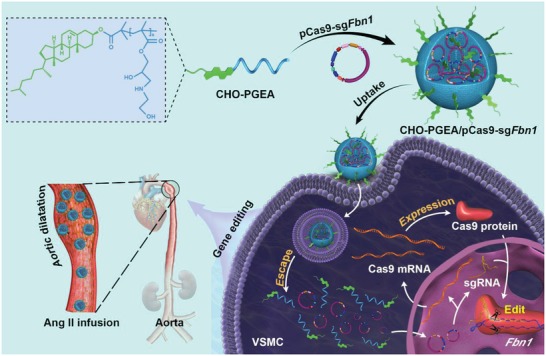
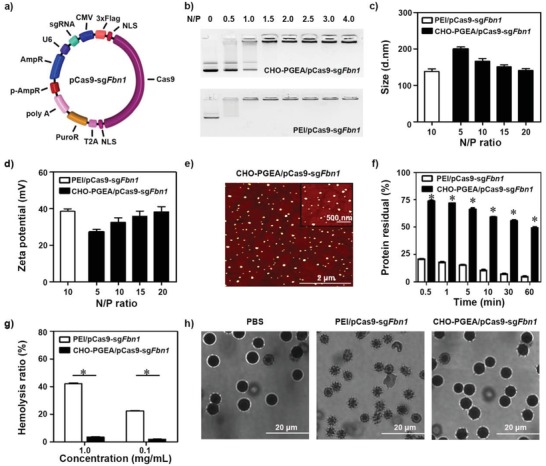
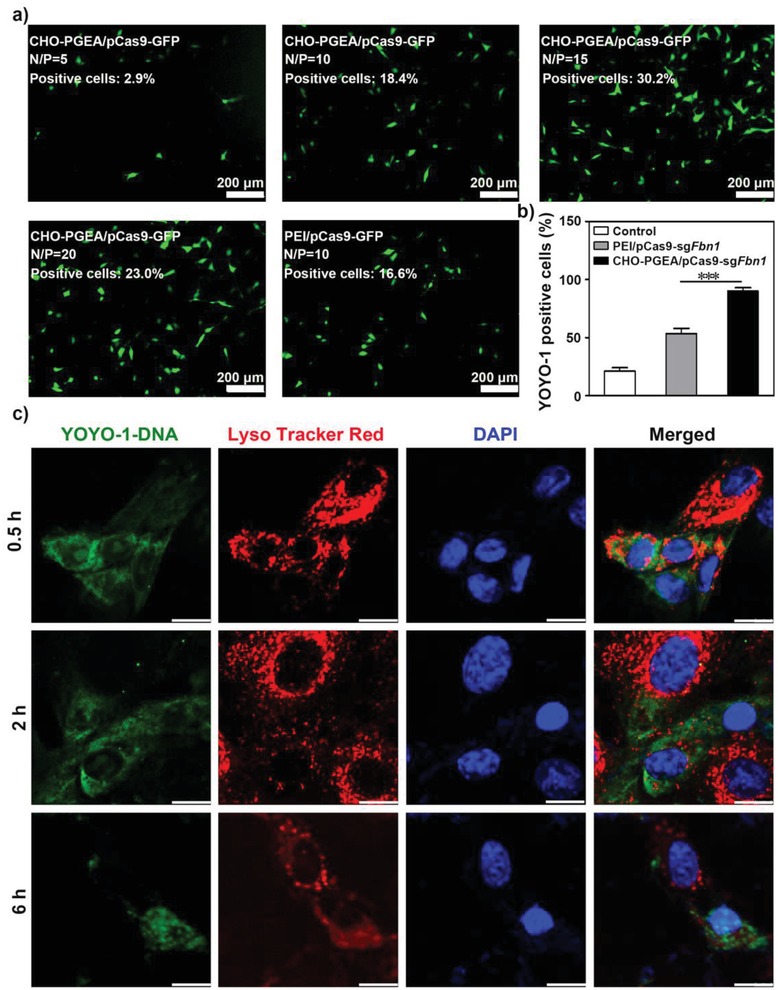
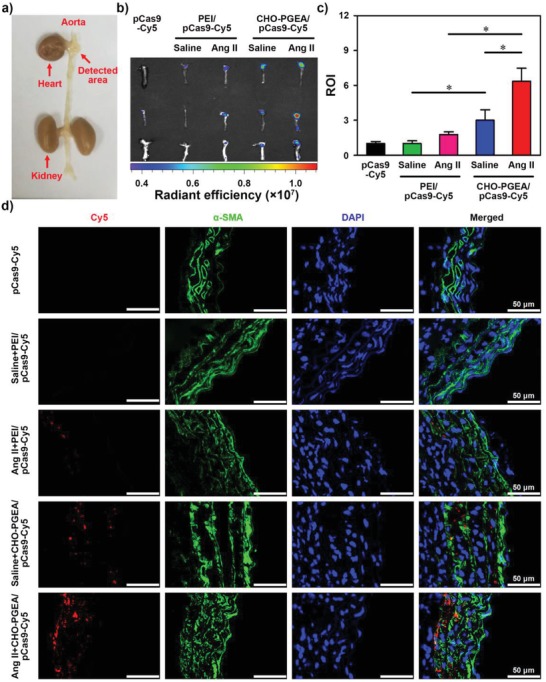
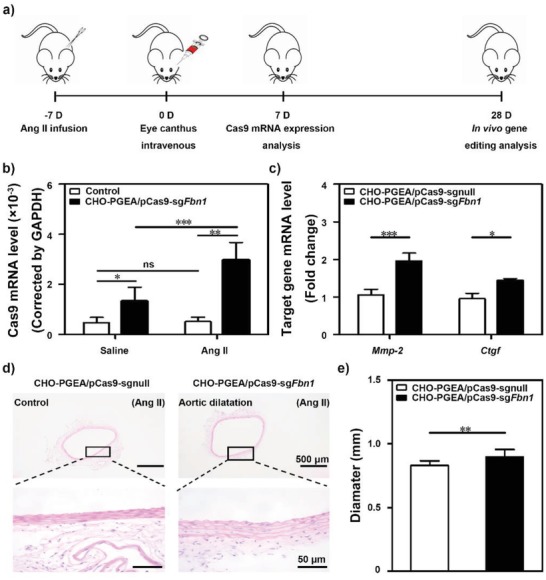
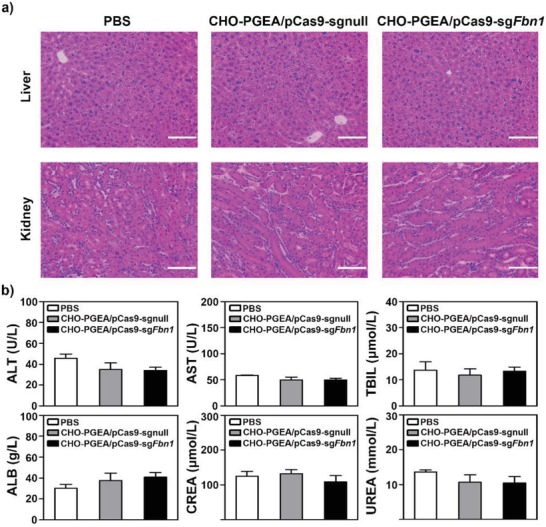
A CRISPR/Cas9 system has emerged as a powerful tool for gene editing to treat genetic mutation related diseases. Due to the complete endothelial barrier, effective delivery of the CRISPR/Cas9 system to vasculatures remains a challenge for in vivo gene editing of genetic vascular diseases especially in aorta. Herein, it is reported that CHO-PGEA (cholesterol (CHO)-terminated ethanolamine-aminated poly(glycidyl methacrylate)) with rich hydroxyl groups can deliver a plasmid based pCas9-sgFbn1 system for the knockout of exon 10 in Fbn1 gene. This is the first report of a polycation-mediated CRISPR/Cas9 system for gene editing in aorta of adult mice. CHO-PGEA/pCas9-sgFbn1 nanosystems can effectively contribute to the knockout of exon 10 in Fbn1 in vascular smooth muscle cells in vitro, which leads to the change of the phosphorylation of Smad2/3 and the increased expression of two downstream signals of Fbn1: Mmp-2 and Ctgf. For in vivo application, the aortic enrichment of CHO-PGEA/Cas9-sgFbn1 is achieved by administering a pressor dose of angiotensin II (Ang II). The effects of the pCas9-sgFbn1 system targeting Fbn1 demonstrate an increase in the expression of Mmp-2 and Ctgf in aorta. Thus, the combination of CHO-PGEA/pCas9-sgFbn1 nanosystems with Ang II infusion can provide the possibility for in vivo gene editing in aorta.
Keywords: CRISPR‐associated nuclease 9 delivery; aorta disease; cationic carriers; genome editing; hydroxyl‐rich.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Polymeric micellar nanoparticles for effective CRISPR/Cas9 genome editing in cancer.Biomaterials. 2024 Sep;309:122573. doi: 10.1016/j.biomaterials.2024.122573. Epub 2024 Apr 17. Biomaterials. 2024. PMID: 38677222
-
Charge-reversal nanocomolexes-based CRISPR/Cas9 delivery system for loss-of-function oncogene editing in hepatocellular carcinoma.J Control Release. 2021 May 10;333:362-373. doi: 10.1016/j.jconrel.2021.03.030. Epub 2021 Mar 27. J Control Release. 2021. PMID: 33785418
-
Effective Delivery of Hypertrophic miRNA Inhibitor by Cholesterol-Containing Nanocarriers for Preventing Pressure Overload Induced Cardiac Hypertrophy.Adv Sci (Weinh). 2019 Apr 6;6(11):1900023. doi: 10.1002/advs.201900023. eCollection 2019 Jun 5. Adv Sci (Weinh). 2019. PMID: 31179215 Free PMC article.
-
Delivery of CRISPR/Cas9 for therapeutic genome editing.J Gene Med. 2019 Jul;21(7):e3107. doi: 10.1002/jgm.3107. J Gene Med. 2019. PMID: 31237055 Review.
-
Therapeutic Genome Editing and In Vivo Delivery.AAPS J. 2021 Jun 2;23(4):80. doi: 10.1208/s12248-021-00613-w. AAPS J. 2021. PMID: 34080099 Review.
Cited by
-
CRISPR/Cas9 therapeutics: progress and prospects.Signal Transduct Target Ther. 2023 Jan 16;8(1):36. doi: 10.1038/s41392-023-01309-7. Signal Transduct Target Ther. 2023. PMID: 36646687 Free PMC article. Review.
-
Advances and challenges in regenerative therapies for abdominal aortic aneurysm.Front Cardiovasc Med. 2024 Jun 4;11:1369785. doi: 10.3389/fcvm.2024.1369785. eCollection 2024. Front Cardiovasc Med. 2024. PMID: 38895536 Free PMC article. Review.
-
Precision in Action: The Role of Clustered Regularly Interspaced Short Palindromic Repeats/Cas in Gene Therapies.Vaccines (Basel). 2024 Jun 7;12(6):636. doi: 10.3390/vaccines12060636. Vaccines (Basel). 2024. PMID: 38932365 Free PMC article. Review.
-
A Lactose-Derived CRISPR/Cas9 Delivery System for Efficient Genome Editing In Vivo to Treat Orthotopic Hepatocellular Carcinoma.Adv Sci (Weinh). 2020 Jul 21;7(17):2001424. doi: 10.1002/advs.202001424. eCollection 2020 Sep. Adv Sci (Weinh). 2020. PMID: 32995132 Free PMC article.
-
Pamoic acid and carbenoxolone specifically inhibit CRISPR/Cas9 in bacteria, mammalian cells, and mice in a DNA topology-specific manner.Genome Biol. 2025 Mar 28;26(1):75. doi: 10.1186/s13059-025-03521-w. Genome Biol. 2025. PMID: 40156040 Free PMC article.
References
-
- Suzuki K., Tsunekawa Y., Hernandez‐Benitez R., Wu J., Zhu J., Kim E. J., Hatanaka F., Yamamoto M., Araoka T., Li Z., Kurita M., Hishida T., Li M., Aizawa E., Guo S., Chen S., Goebl A., Soligalla R. D., Qu J., Jiang T., Fu X., Jafari M., Esteban C. R., Berggren W. T., Lajara J., Nunez‐Delicado E., Guillen P., Campistol J. M., Matsuzaki F., Liu G. H., Magistretti P., Zhang K., Callaway E. M., Zhang K., Belmonte J. C., Nature 2016, 540, 144. - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous