

The Interaction of Klebsiella pneumoniae With Lipid Rafts-Associated Cholesterol Increases Macrophage-Mediated Phagocytosis Due to Down Regulation of the Capsule Polysaccharide
- PMID: 31380298
- PMCID: PMC6650577
- DOI: 10.3389/fcimb.2019.00255
The Interaction of Klebsiella pneumoniae With Lipid Rafts-Associated Cholesterol Increases Macrophage-Mediated Phagocytosis Due to Down Regulation of the Capsule Polysaccharide
Abstract
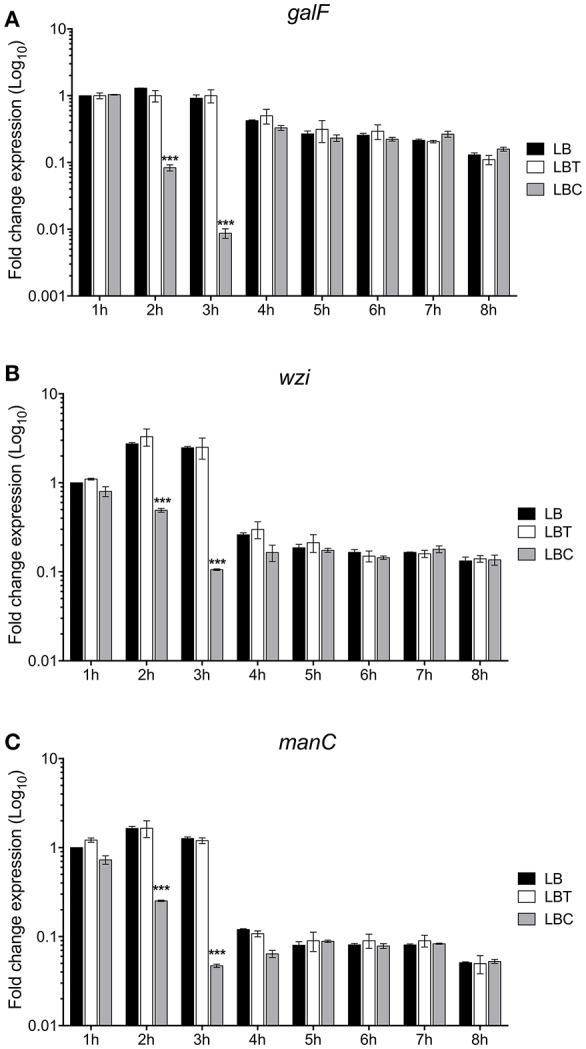
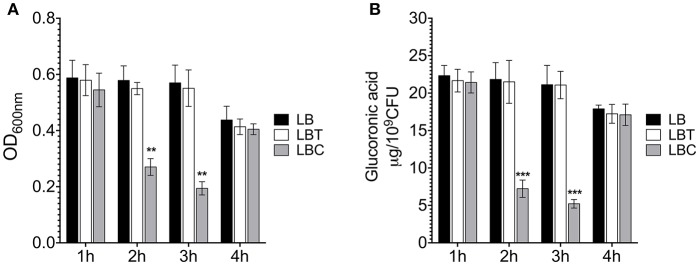
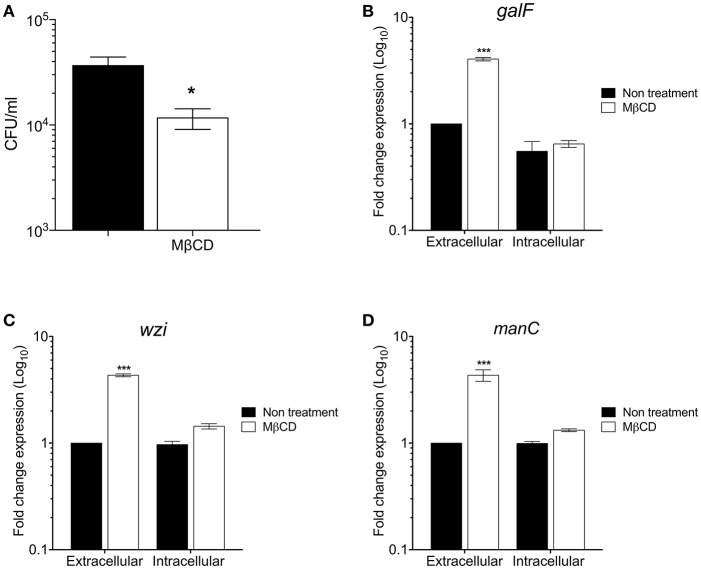
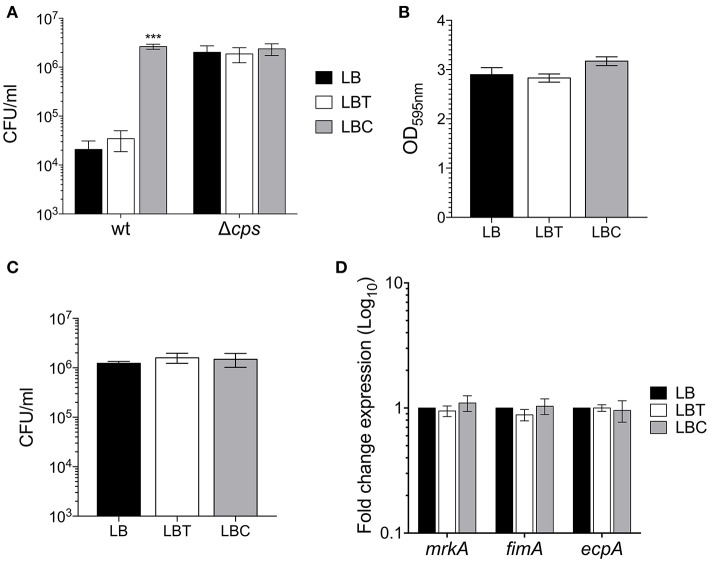
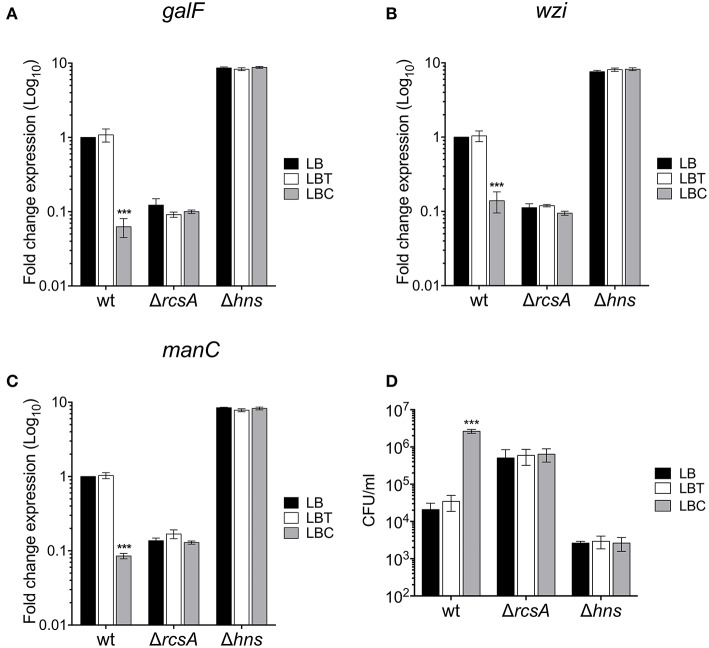
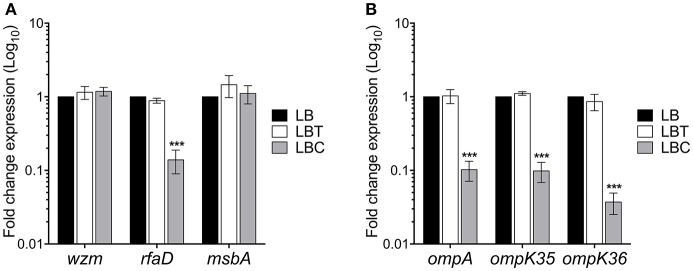
Klebsiella pneumoniae successfully colonizes host tissues by recognizing and interacting with cholesterol present on membrane-associated lipid rafts. In this study, we evaluated the role of cholesterol in the expression of capsule polysaccharide genes of K. pneumoniae and its implication in resistance to phagocytosis. Our data revealed that exogenous cholesterol added to K. pneumoniae increases macrophage-mediated phagocytosis. To explain this event, the expression of capsular galF, wzi, and manC genes was determined in the presence of cholesterol. Down-regulation of these capsular genes occurred leading to increased susceptibility to phagocytosis by macrophages. In contrast, depletion of cholesterol from macrophage membranes led to enhanced expression of galF, wzi, and manC genes and to capsule production resulting in resistance to macrophage-mediated phagocytosis. Cholesterol-mediated repression of capsular genes was dependent on the RcsA and H-NS global regulators. Finally, cholesterol also down-regulated the expression of genes responsible for LPS core oligosaccharides production and OMPs. Our results suggest that cholesterol plays an important role for the host by reducing the anti-phagocytic properties of the K. pneumoniae capsule facilitating bacterial engulfment by macrophages during the bacteria-eukaryotic cell interaction mediated by lipid rafts.
Keywords: H-NS; Klebsiella pneumoniae; RcsA; capsule; cholesterol; phagocytosis.
Figures
References
-
- Ares M. A., Alcantar-Curiel M. D., Jimenez-Galicia C., Rios-Sarabia N., Pacheco S., De la Cruz M. A. (2013). Antibiotic resistance of gram-negative bacilli isolated from pediatric patients with nosocomial bloodstream infections in a Mexican tertiary care hospital. Chemotherapy 59, 361–368. 10.1159/000362085 - DOI - PubMed
-
- Ares M. A., Fernandez-Vazquez J. L., Rosales-Reyes R., Jarillo-Quijada M. D., Von Bargen K., Torres J., et al. (2016). H-NS nucleoid protein controls virulence features of Klebsiella pneumoniae by regulating the expression of type 3 pili and the capsule polysaccharide. Front. Cell Infect. Microbiol. 6:13. 10.3389/fcimb.2016.00013 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous