

DPP8/9 inhibitors are universal activators of functional NLRP1 alleles
- PMID: 31383852
- PMCID: PMC6683174
- DOI: 10.1038/s41419-019-1817-5
DPP8/9 inhibitors are universal activators of functional NLRP1 alleles
Abstract
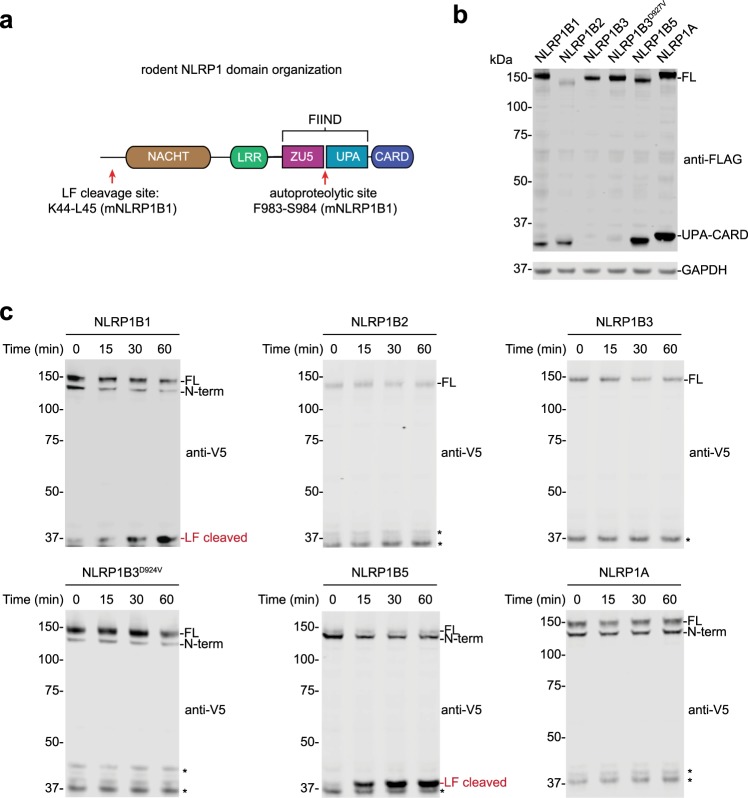
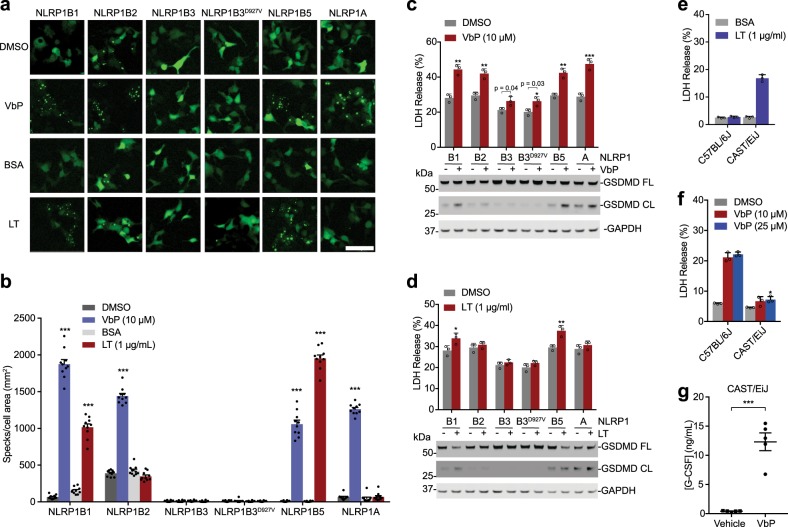
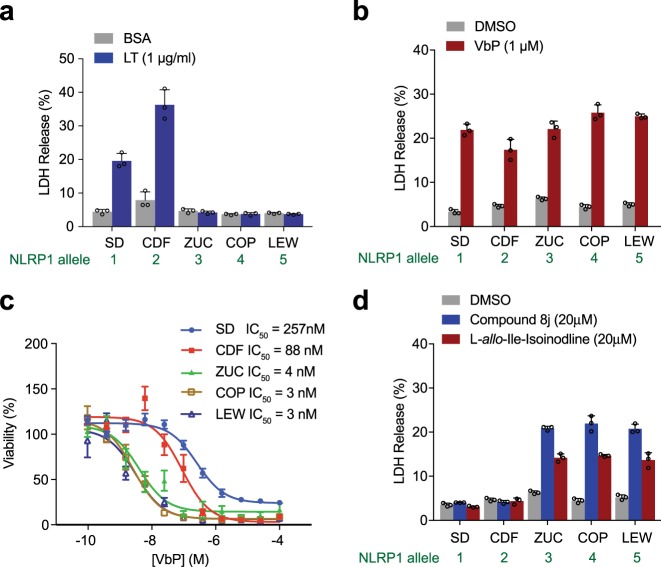
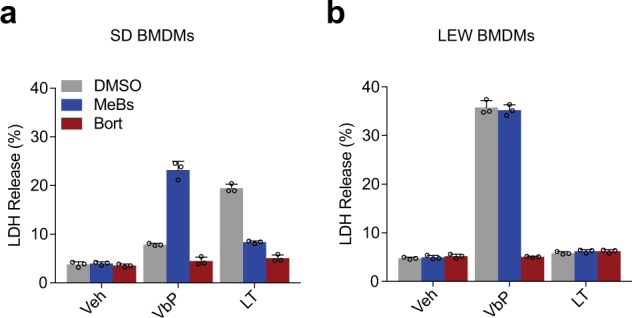
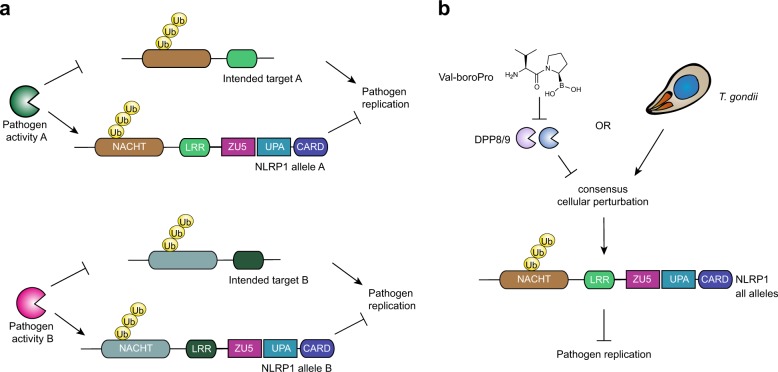
Intracellular pathogenic structures or activities stimulate the formation of inflammasomes, which recruit and activate caspase-1 and trigger an inflammatory form of cell death called pyroptosis. The well-characterized mammalian inflammasome sensor proteins all detect one specific type of signal, for example double-stranded DNA or bacterial flagellin. Remarkably, NLRP1 was the first protein discovered to form an inflammasome, but the pathogenic signal that NLRP1 detects has not yet been identified. NLRP1 is highly polymorphic, even among inbred rodent strains, and it has been suggested that these diverse NLRP1 alleles may have evolved to detect entirely different stimuli. Intriguingly, inhibitors of the serine proteases DPP8 and DPP9 (DPP8/9) were recently shown to activate human NLRP1, its homolog CARD8, and several mouse NLRP1 alleles. Here, we show now that DPP8/9 inhibitors activate all functional rodent NLRP1 alleles, indicating that DPP8/9 inhibition induces a signal detected by all NLRP1 proteins. Moreover, we discovered that the NLRP1 allele sensitivities to DPP8/9 inhibitor-induced and Toxoplasma gondii-induced pyroptosis are strikingly similar, suggesting that DPP8/9 inhibition phenocopies a key activity of T. gondii. Overall, this work indicates that the highly polymorphic NLRP1 inflammasome indeed senses a specific signal like the other mammalian inflammasomes.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
