

Tundra microbial community taxa and traits predict decomposition parameters of stable, old soil organic carbon
- PMID: 31384013
- PMCID: PMC6863828
- DOI: 10.1038/s41396-019-0485-x
Tundra microbial community taxa and traits predict decomposition parameters of stable, old soil organic carbon
Abstract
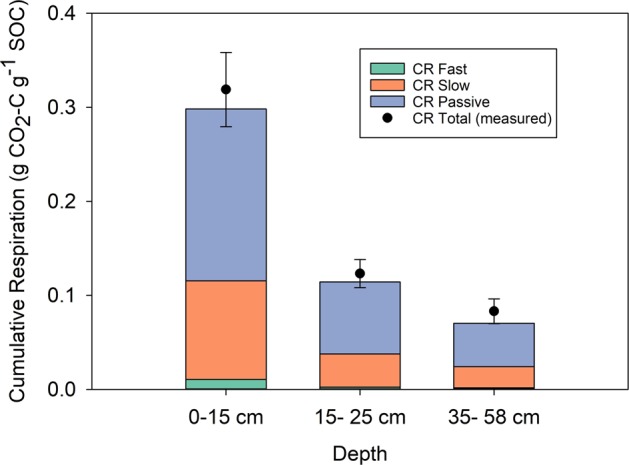
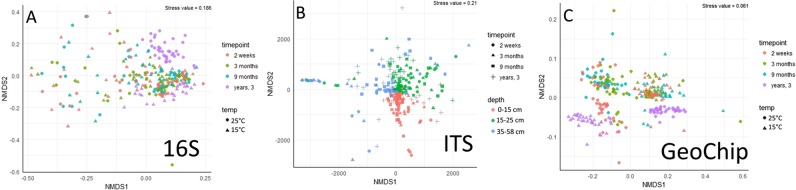
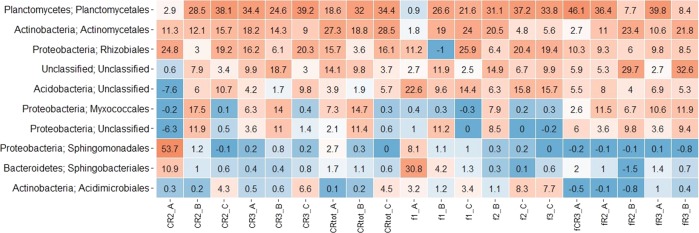
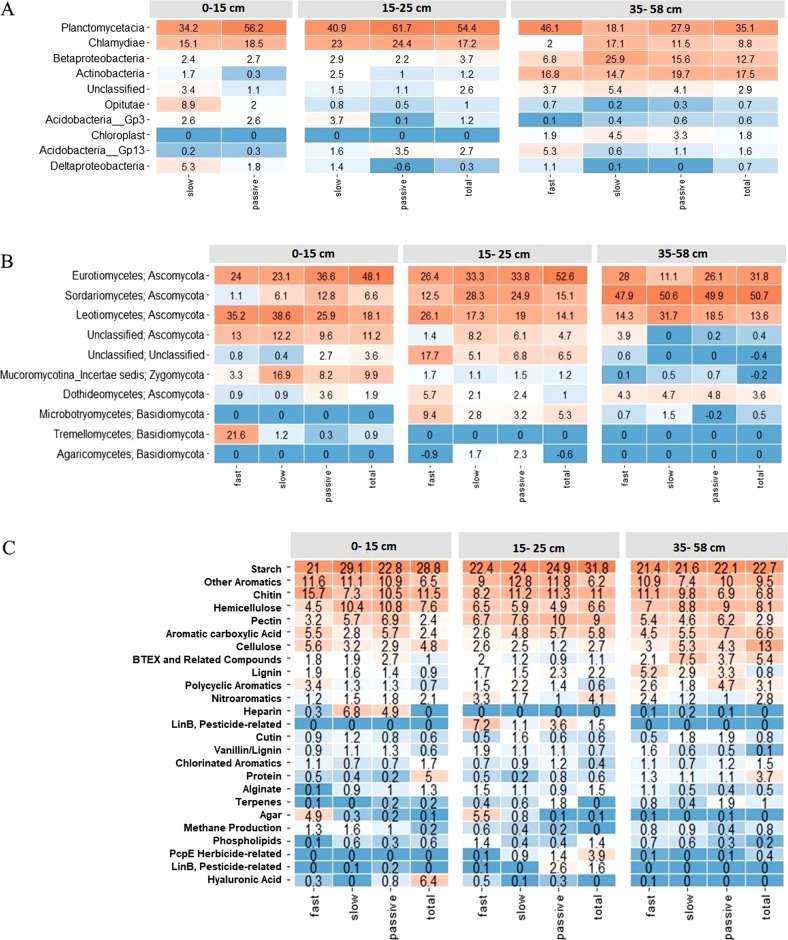
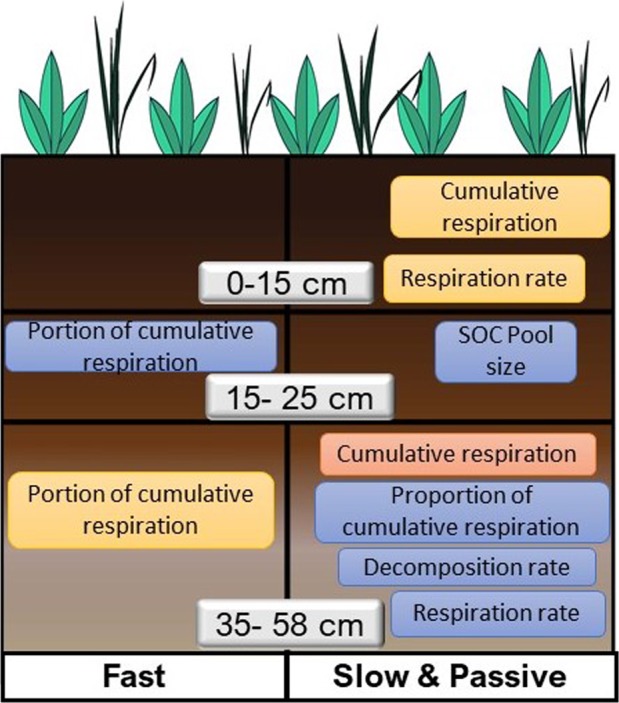
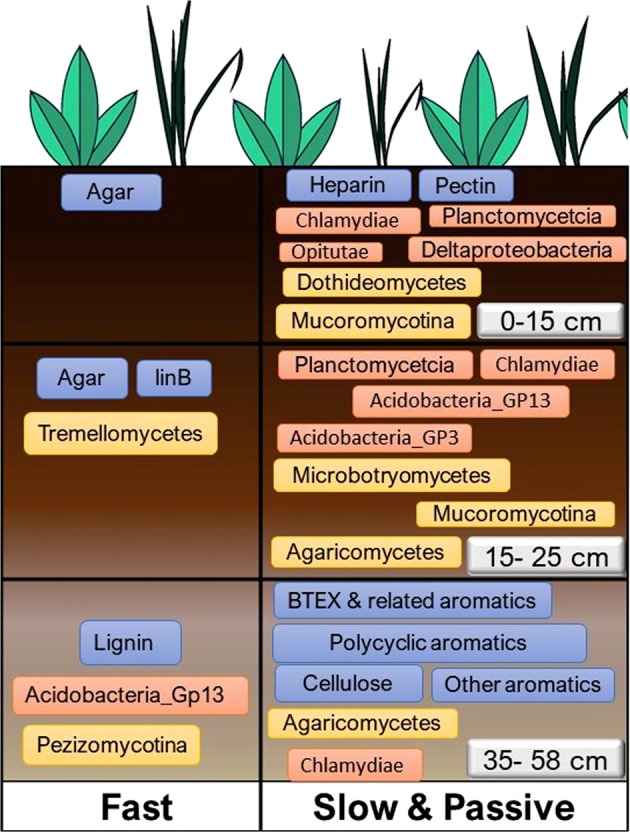
The susceptibility of soil organic carbon (SOC) in tundra to microbial decomposition under warmer climate scenarios potentially threatens a massive positive feedback to climate change, but the underlying mechanisms of stable SOC decomposition remain elusive. Herein, Alaskan tundra soils from three depths (a fibric O horizon with litter and course roots, an O horizon with decomposing litter and roots, and a mineral-organic mix, laying just above the permafrost) were incubated. Resulting respiration data were assimilated into a 3-pool model to derive decomposition kinetic parameters for fast, slow, and passive SOC pools. Bacterial, archaeal, and fungal taxa and microbial functional genes were profiled throughout the 3-year incubation. Correlation analyses and a Random Forest approach revealed associations between model parameters and microbial community profiles, taxa, and traits. There were more associations between the microbial community data and the SOC decomposition parameters of slow and passive SOC pools than those of the fast SOC pool. Also, microbial community profiles were better predictors of model parameters in deeper soils, which had higher mineral contents and relatively greater quantities of old SOC than in surface soils. Overall, our analyses revealed the functional potential of microbial communities to decompose tundra SOC through a suite of specialized genes and taxa. These results portray divergent strategies by which microbial communities access SOC pools across varying depths, lending mechanistic insights into the vulnerability of what is considered stable SOC in tundra regions.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Koven CD, Riley WJ, Stern A. Analysis of permafrost thermal dynamics and response to climate change in the CMIP5 Earth System Models. J Clim. 2013;26:1877–900. doi: 10.1175/JCLI-D-12-00228.1. - DOI
-
- Schuur E, Abbott B, Bowden W, Brovkin V, Camill P, Canadell J, et al. Expert assessment of vulnerability of permafrost carbon to climate change. Clim Change. 2013;119:359–74. doi: 10.1007/s10584-013-0730-7. - DOI
-
- Schuur EA, Bockheim J, Canadell JG, Euskirchen E, Field CB, Goryachkin SV, et al. Vulnerability of permafrost carbon to climate change: Implications for the global carbon cycle. AIBS Bull. 2008;58:701–14.
