

Long-term repopulation of aged bone marrow stem cells using young Sca-1 cells promotes aged heart rejuvenation
- PMID: 31385396
- PMCID: PMC6826122
- DOI: 10.1111/acel.13026
Long-term repopulation of aged bone marrow stem cells using young Sca-1 cells promotes aged heart rejuvenation
Abstract
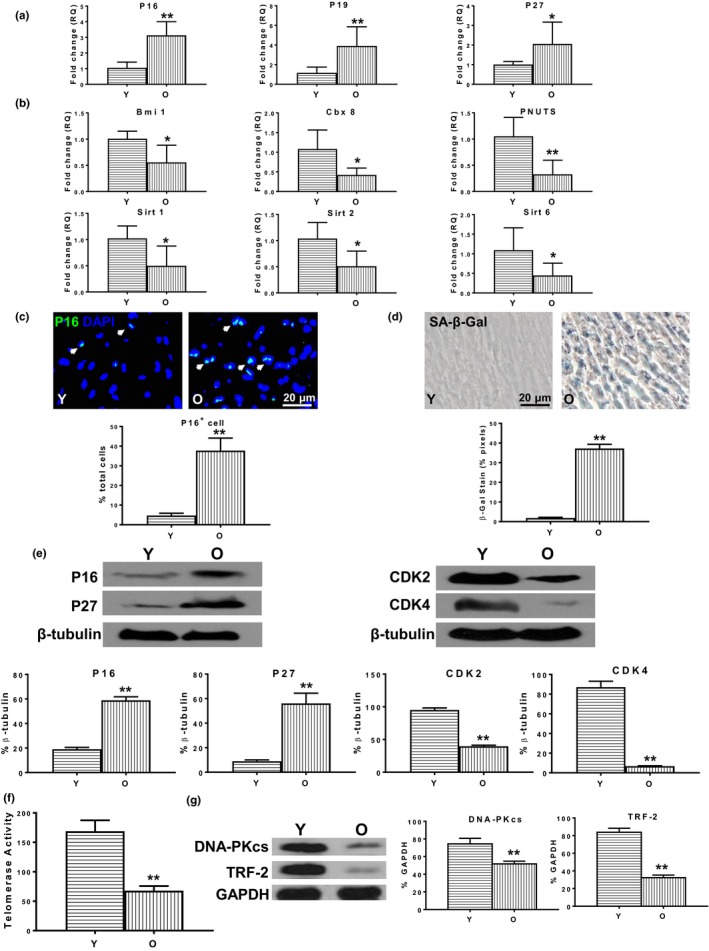
Reduced quantity and quality of stem cells in aged individuals hinders cardiac repair and regeneration after injury. We used young bone marrow (BM) stem cell antigen 1 (Sca-1) cells to reconstitute aged BM and rejuvenate the aged heart, and examined the underlying molecular mechanisms. BM Sca-1+ or Sca-1- cells from young (2-3 months) or aged (18-19 months) GFP transgenic mice were transplanted into lethally irradiated aged mice to generate 4 groups of chimeras: young Sca-1+ , young Sca-1- , old Sca-1+ , and old Sca-1- . Four months later, expression of rejuvenation-related genes (Bmi1, Cbx8, PNUTS, Sirt1, Sirt2, Sirt6) and proteins (CDK2, CDK4) was increased along with telomerase activity and telomerase-related protein (DNA-PKcs, TRF-2) expression, whereas expression of senescence-related genes (p16INK4a , P19ARF , p27Kip1 ) and proteins (p16INK4a , p27Kip1 ) was decreased in Sca-1+ chimeric hearts, especially in the young group. Host cardiac endothelial cells (GFP- CD31+ ) but not cardiomyocytes were the primary cell type rejuvenated by young Sca-1+ cells as shown by improved proliferation, migration, and tubular formation abilities. C-X-C chemokine CXCL12 was the factor most highly expressed in homed donor BM (GFP+ ) cells isolated from young Sca-1+ chimeric hearts. Protein expression of Cxcr4, phospho-Akt, and phospho-FoxO3a in endothelial cells derived from the aged chimeric heart was increased, especially in the young Sca-1+ group. Reconstitution of aged BM with young Sca-1+ cells resulted in effective homing of functional stem cells in the aged heart. These young, regenerative stem cells promoted aged heart rejuvenation through activation of the Cxcl12/Cxcr4 pathway of cardiac endothelial cells.
Keywords: Sca-1; aging; heart; reconstitution; rejuvenation; stem cells.
© 2019 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures





References
-
- Brunet, A. , Bonni, A. , Zigmond, M. J. , Lin, M. Z. , Juo, P. , Hu, L. S. , … Greenberg, M. E. (1999). Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell, 96(6), 857–868. - PubMed
-
- Chen, S. , Lin, J. , Matsuguchi, T. , Blackburn, E. , Yeh, F. , Best, L. G. , … Zhao, J. (2014). Short leukocyte telomere length predicts incidence and progression of carotid atherosclerosis in American Indians: The Strong Heart Family Study. Aging (Albany NY), 6(5), 414–427. 10.18632/aging.100671 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1201581618/Bureau of Education of Guangzhou Municipality/International
- RE07-010/Ontario Research Fund/International
- 332652/CIHR/Canada
- 2016A020215166/Science and Technology Planning Project of Guangdong Province/International
- 2017A020215084/Science and Technology Planning Project of Guangdong Province/International
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
