

Subterranean Termite Social Alarm and Hygienic Responses to Fungal Pathogens
- PMID: 31387197
- PMCID: PMC6723859
- DOI: 10.3390/insects10080240
Subterranean Termite Social Alarm and Hygienic Responses to Fungal Pathogens
Abstract
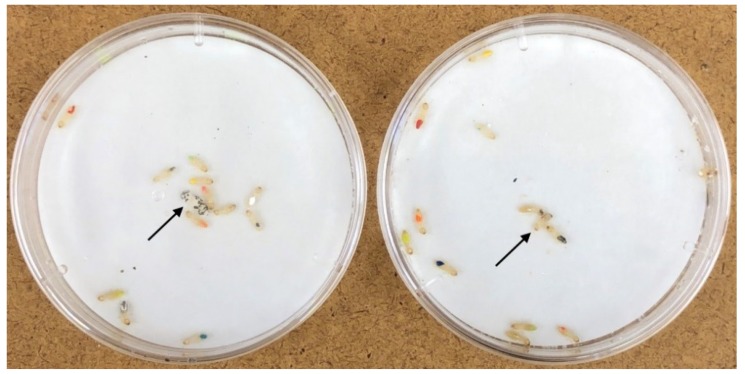
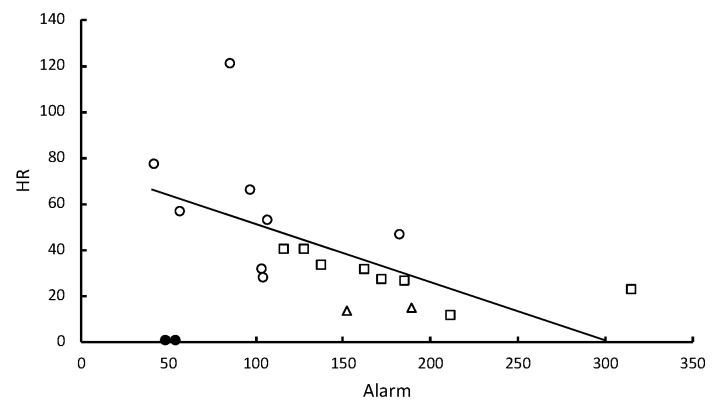
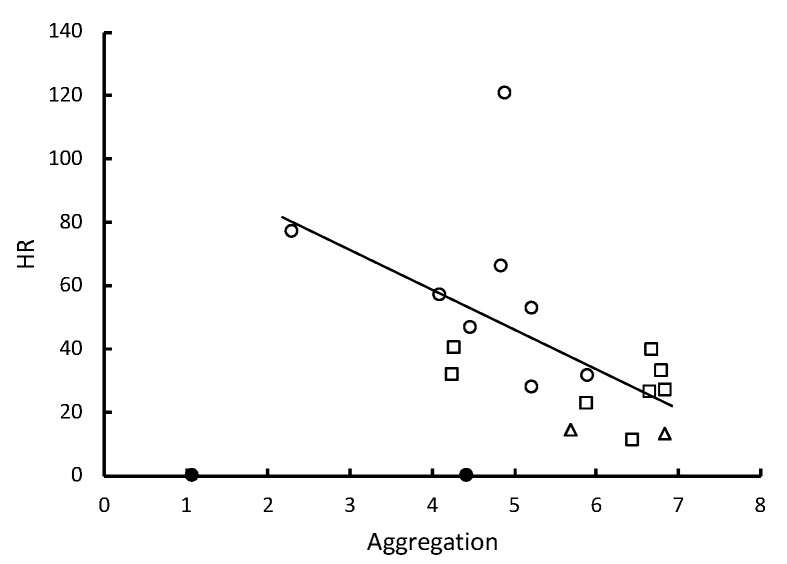
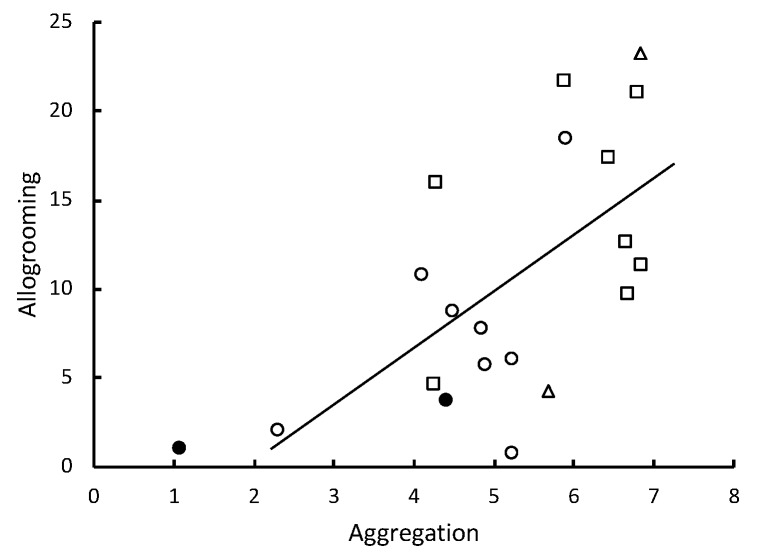
In social insects, alerting nestmates to the presence of a pathogen should be critical for limiting its spread and initiating social mechanisms of defense. Here we show that subterranean termites use elevated vibratory alarm behavior to help prevent fatal fungal infections. The elevated alarm leads to elevated social hygiene. This requires that termites coalesce so that they can groom each other's cuticular surfaces of contaminating conidial spores. Groups of 12 Reticulitermes flavipes workers varied in their response when immersed in conidia solutions of nine different strains of Metarhizium. Pathogen alarm displays of short 2-7-second bursts of rapid longitudinal oscillatory movement (LOM), observed over 12 min following a fungal challenge, were positively correlated with the time that workers spent aggregated together grooming each other. The frequency of these LOMs was inversely correlated with fatal fungal infections. The variation in fatalities appeared to be largely attributable to a differential response to Metarhizium brunneum and Metarhizium robertsii in the time spent in aggregations and the frequency of allogrooming. Isolated workers challenged with conidia did not display LOMs, which suggests that the alarm is a conditional social response. LOMs appear to help signal the presence of fungal pathogens whose virulence depends on the level of this emergency alert.
Keywords: Metarhizium; alarm behavior; allogrooming; entomopathogens; social immunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Rosengaus R.B., Traniello J.F., Bulmer M.S. Ecology, behavior and evolution of disease resistance in termites. In: Bignell D.E., Roisin Y., Lo N., editors. Biology of Termites: A Modern Synthesis. Springer; London, UK: 2011. pp. 165–192.
-
- Yanagawa A., Shimizu S. Resistance of the termite, Coptotermes formosanus Shiraki to Metarhizium anisopliae due to grooming. BioControl. 2007;52:75–85. doi: 10.1007/s10526-006-9020-x. - DOI
-
- Tranter C., Hughes W. Acid, silk and grooming: Alternative strategies in social immunity in ants? Behav. Ecol. Sociobiol. 2015;69:1687–1699. doi: 10.1007/s00265-015-1980-3. - DOI
-
- Hajek A.E. Ecology of terrestrial fungal entomopathogens. Adv. Microb. Ecol. 1997;15:193–249.
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
