

Thymic Stromal Lymphopoietin Interferes with the Apoptosis of Human Skin Mast Cells by a Dual Strategy Involving STAT5/Mcl-1 and JNK/Bcl-xL
- PMID: 31387206
- PMCID: PMC6721763
- DOI: 10.3390/cells8080829
Thymic Stromal Lymphopoietin Interferes with the Apoptosis of Human Skin Mast Cells by a Dual Strategy Involving STAT5/Mcl-1 and JNK/Bcl-xL
Erratum in
-
Correction: Hazzan et al. Thymic Stromal Lymphopoietin Interferes with the Apoptosis of Human Skin Mast Cells by a Dual Strategy Involving STAT5/Mcl-1 and JNK/Bcl-xL. Cells 2019, 8, 829.Cells. 2024 Dec 19;13(24):2105. doi: 10.3390/cells13242105. Cells. 2024. PMID: 39768229 Free PMC article.
Abstract
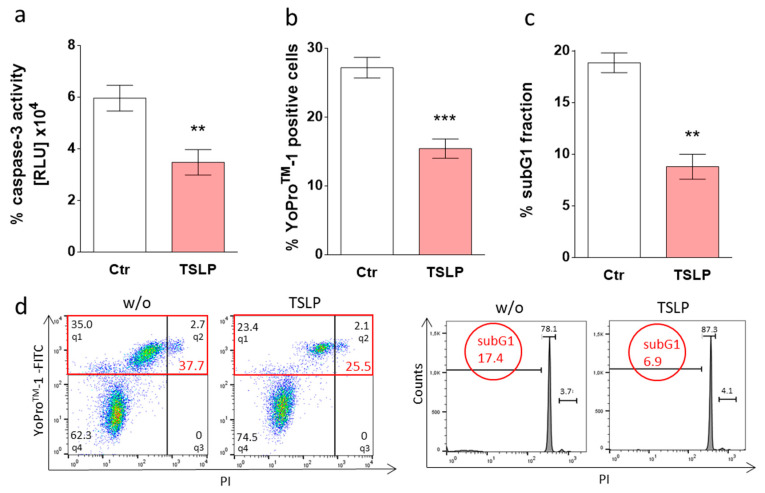
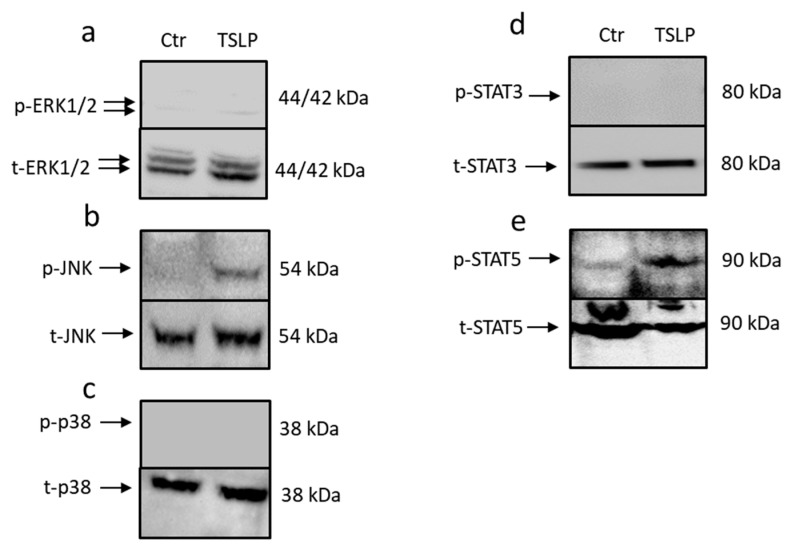
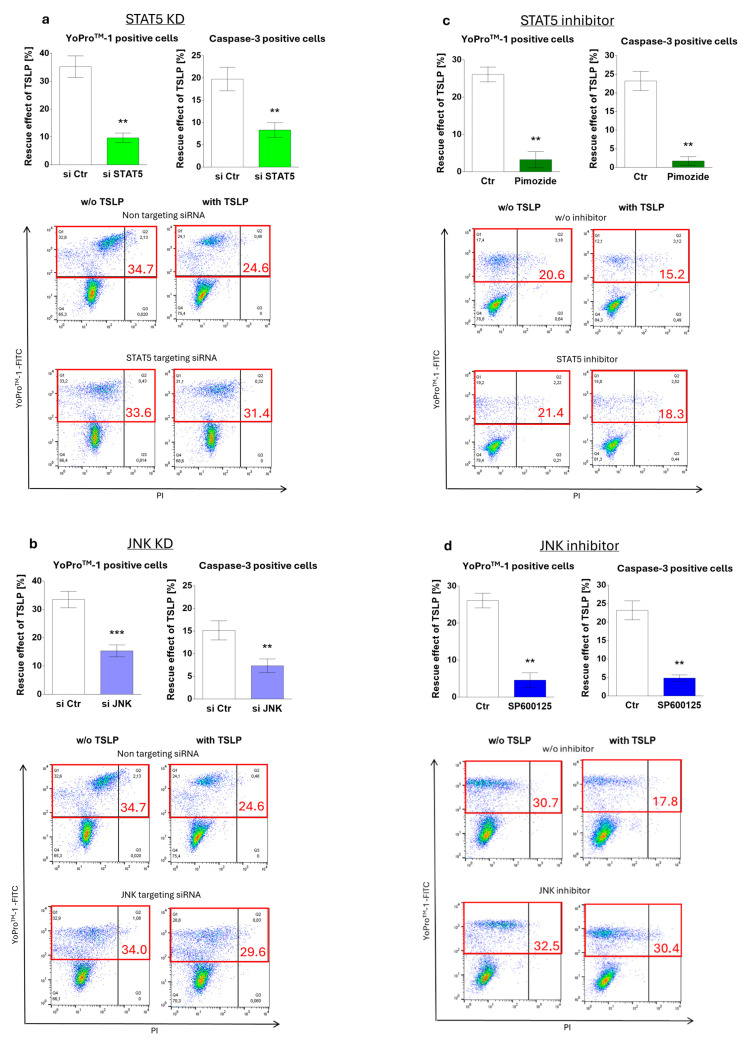
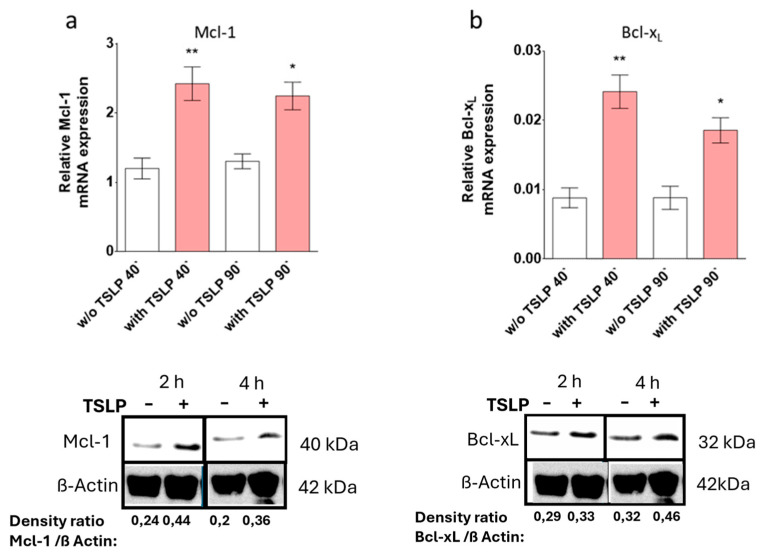
Mast cells (MCs) play critical roles in allergic and inflammatory reactions and contribute to multiple pathologies in the skin, in which they show increased numbers, which frequently correlates with severity. It remains ill-defined how MC accumulation is established by the cutaneous microenvironment, in part because research on human MCs rarely employs MCs matured in the tissue, and extrapolations from other MC subsets have limitations, considering the high level of MC heterogeneity. Thymic stromal lymphopoietin (TSLP)-released by epithelial cells, like keratinocytes, following disturbed homeostasis and inflammation-has attracted much attention, but its impact on skin MCs remains undefined, despite the vast expression of the TSLP receptor by these cells. Using several methods, each detecting a distinct component of the apoptotic process (membrane alterations, DNA degradation, and caspase-3 activity), our study pinpoints TSLP as a novel survival factor of dermal MCs. TSLP confers apoptosis resistance via concomitant activation of the TSLP/ signal transducer and activator of transcription (STAT)-5 / myeloid cell leukemia (Mcl)-1 route and a newly uncovered TSLP/ c-Jun-N-terminal kinase (JNK)/ B-cell lymphoma (Bcl)-xL axis, as evidenced by RNA interference and pharmacological inhibition. Our findings highlight the potential contribution of TSLP to the MC supportive niche of the skin and, vice versa, highlight MCs as crucial responders to TSLP in the context of TSLP-driven disorders.
Keywords: Bcl-xL; JNK; Mcl-1; RNA interference; STAT5; TSLP; apoptosis; mast cells; skin; survival.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
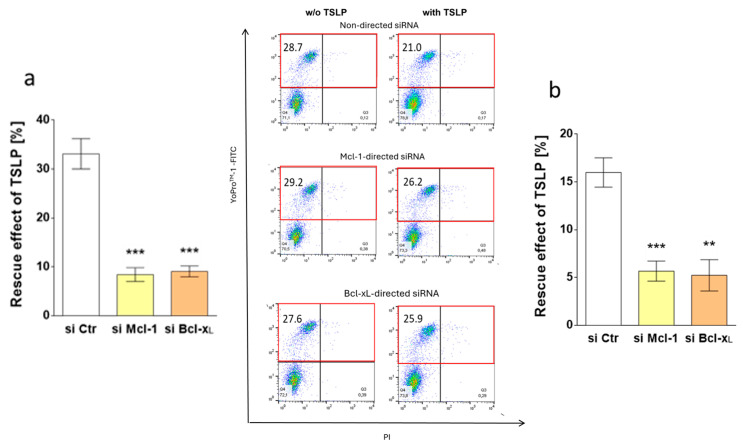
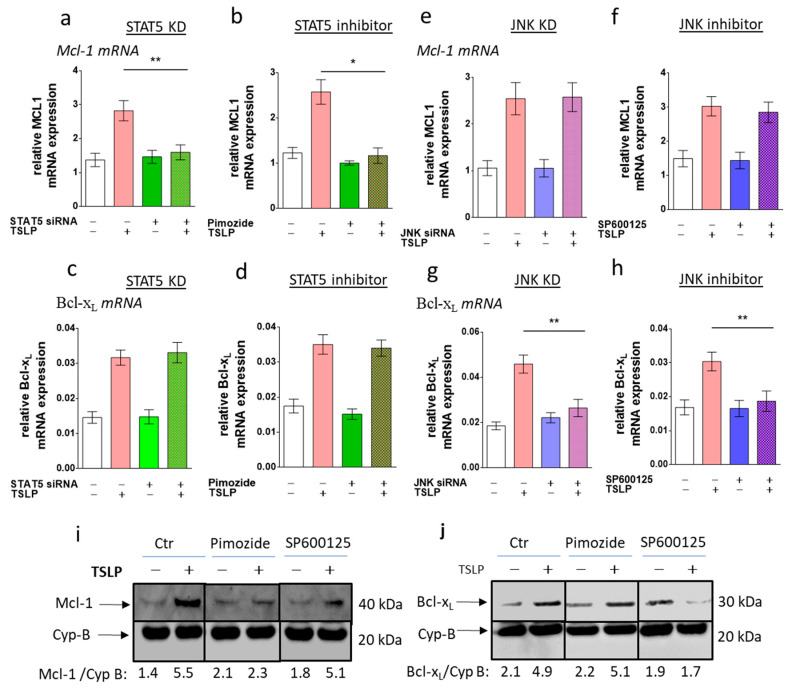
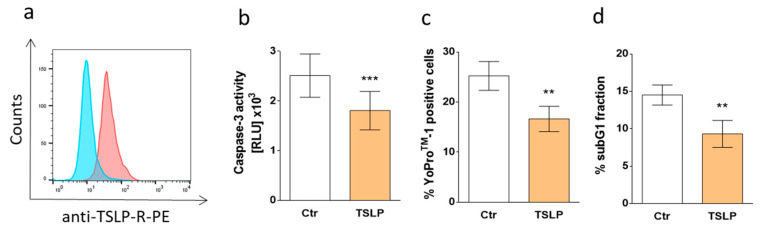
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
