

Transcriptomic Signatures of Postnatal and Adult Intrinsically Photosensitive Ganglion Cells
- PMID: 31387875
- PMCID: PMC6712207
- DOI: 10.1523/ENEURO.0022-19.2019
Transcriptomic Signatures of Postnatal and Adult Intrinsically Photosensitive Ganglion Cells
Abstract
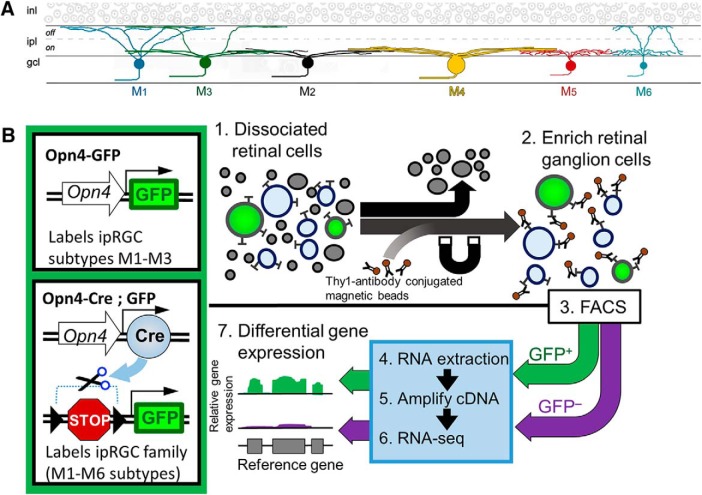
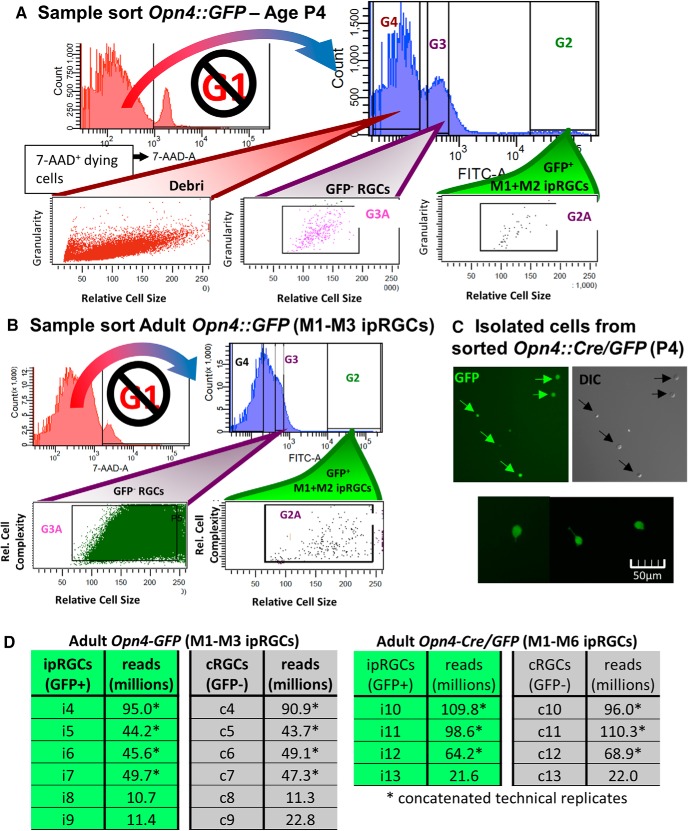
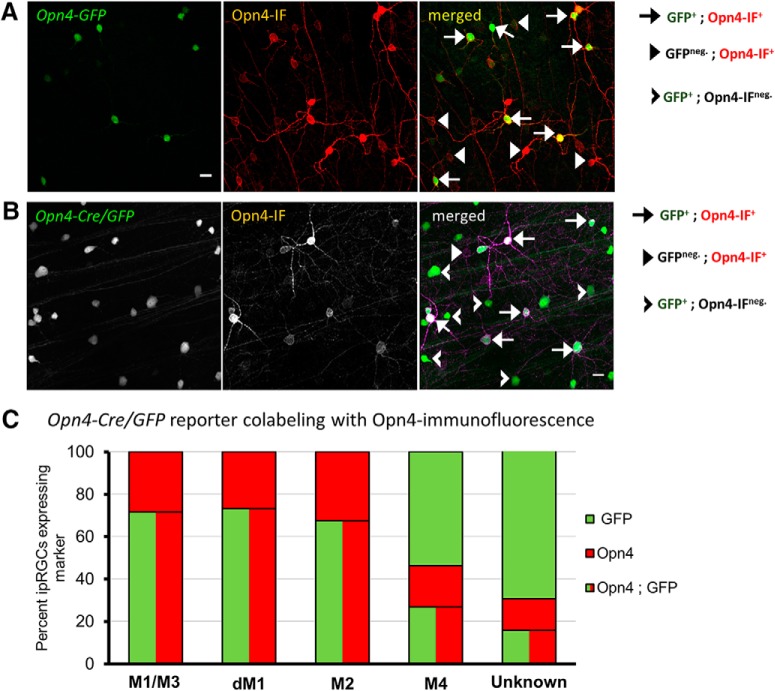
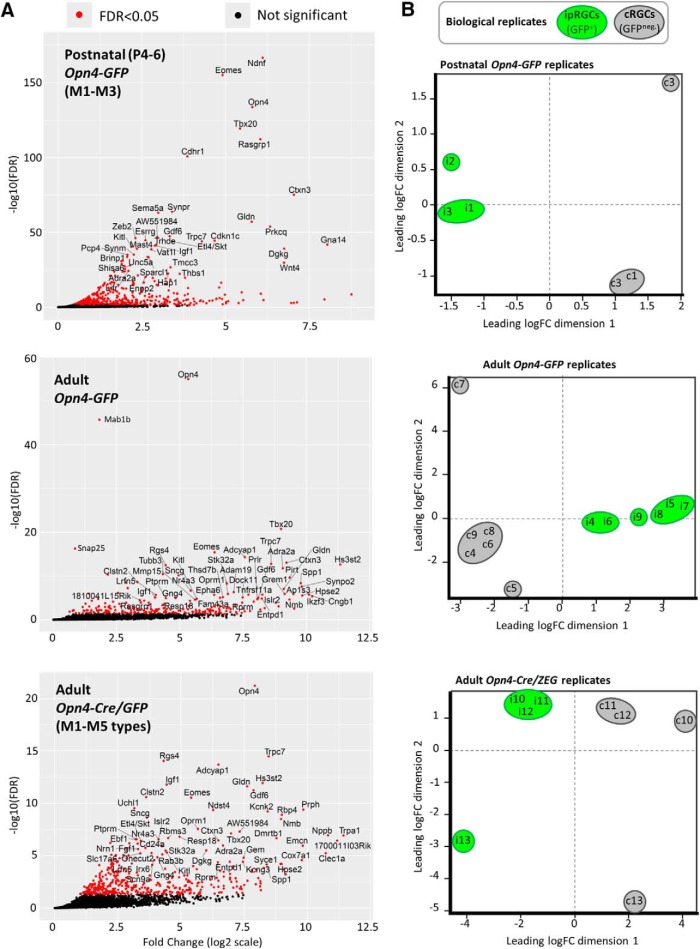
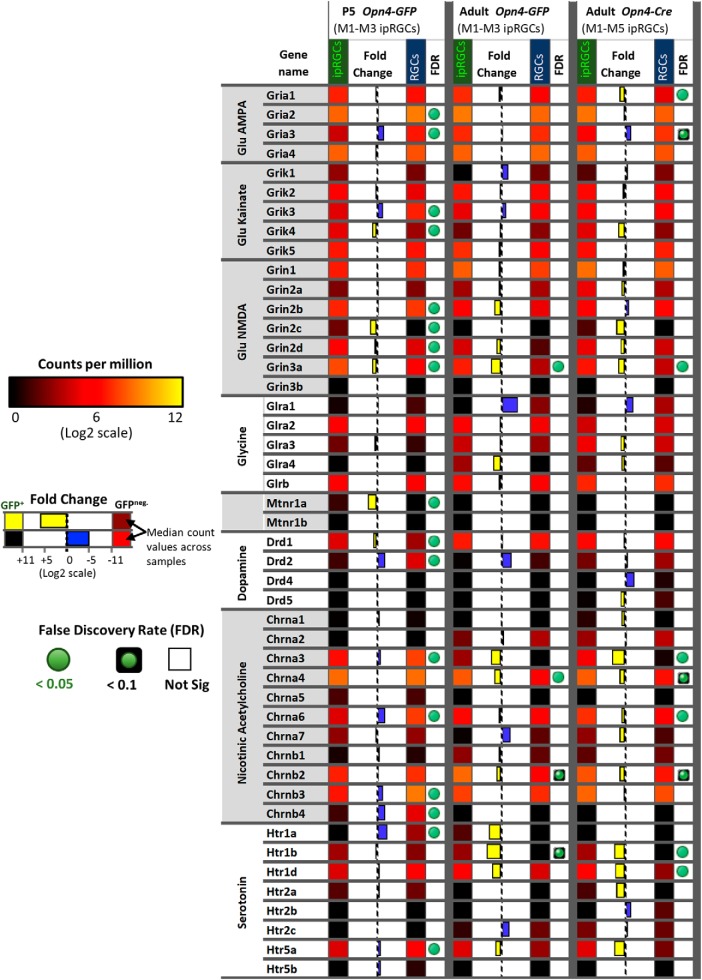
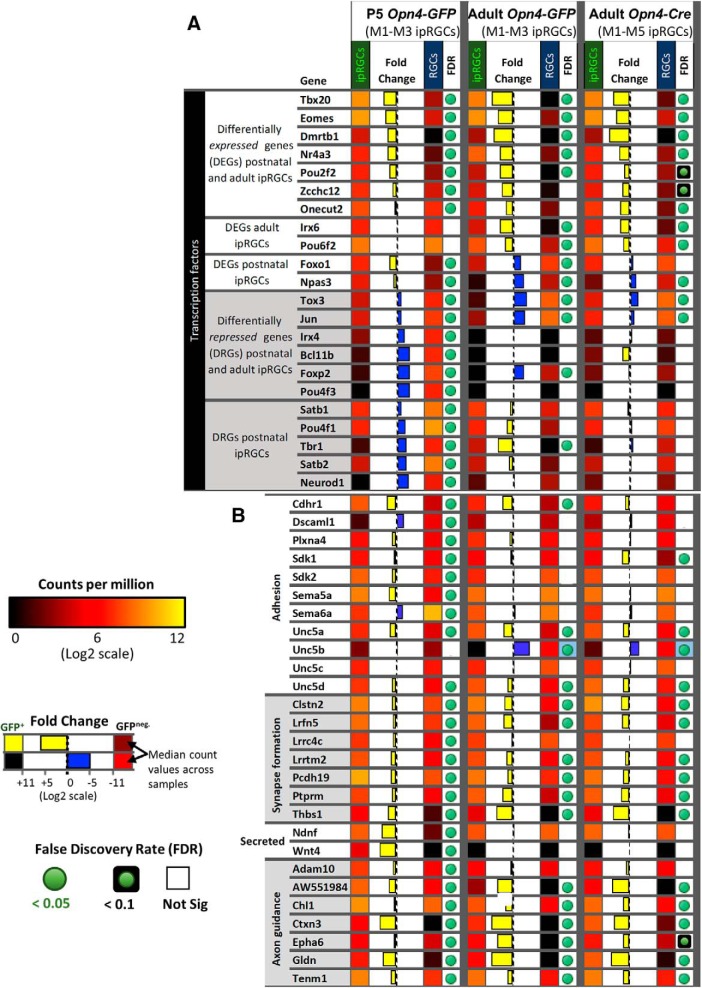
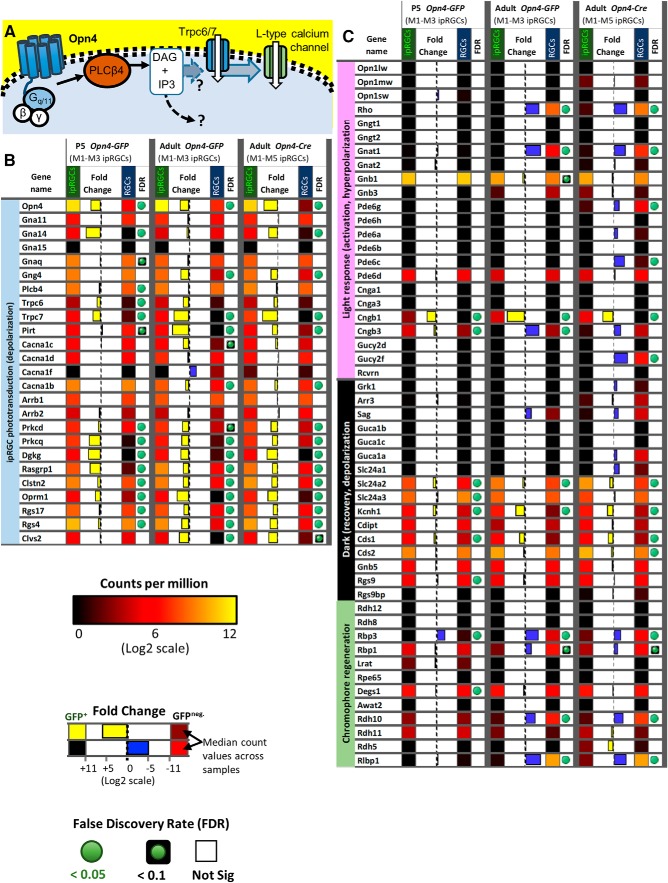
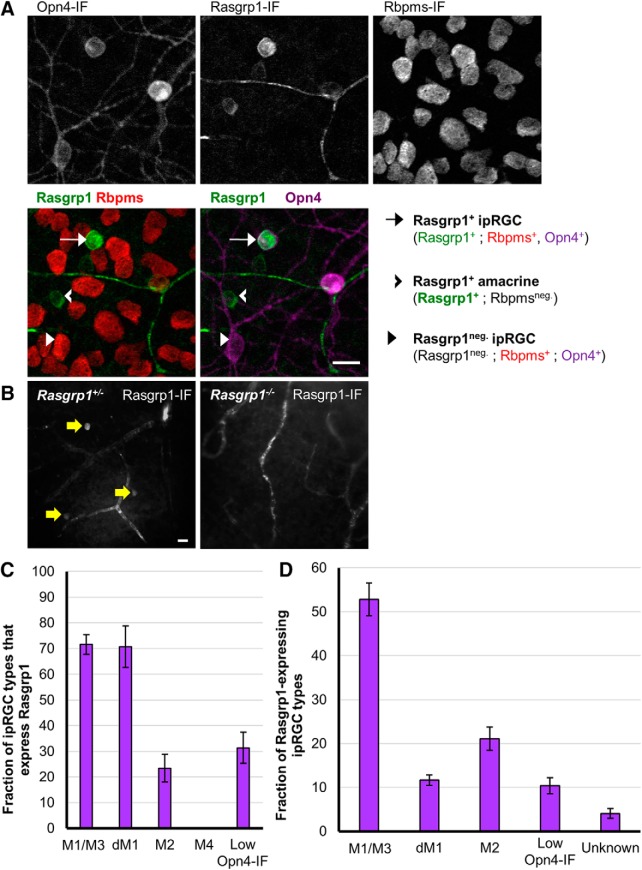
Intrinsically photosensitive retinal ganglion cells (ipRGCs) are rare mammalian photoreceptors essential for non-image-forming vision functions, such as circadian photoentrainment and the pupillary light reflex. They comprise multiple subtypes distinguishable by morphology, physiology, projections, and levels of expression of melanopsin (Opn4), their photopigment. The molecular programs that distinguish ipRGCs from other ganglion cells and ipRGC subtypes from one another remain elusive. Here, we present comprehensive gene expression profiles of early postnatal and adult mouse ipRGCs purified from two lines of reporter mice that mark different sets of ipRGC subtypes. We find dozens of novel genes highly enriched in ipRGCs. We reveal that Rasgrp1 and Tbx20 are selectively expressed in subsets of ipRGCs, though these molecularly defined groups imperfectly match established ipRGC subtypes. We demonstrate that the ipRGCs regulating circadian photoentrainment are diverse at the molecular level. Our findings reveal unexpected complexity in gene expression patterns across mammalian ipRGC subtypes.
Keywords: RNA-seq; cell type; circadian; intrinsically photosensitive; retinal ganglion; vision.
Copyright © 2019 Berg et al.
Figures



Similar articles
-
Degeneration of ipRGCs in Mouse Models of Huntington's Disease Disrupts Non-Image-Forming Behaviors Before Motor Impairment.J Neurosci. 2019 Feb 20;39(8):1505-1524. doi: 10.1523/JNEUROSCI.0571-18.2018. Epub 2018 Dec 26. J Neurosci. 2019. PMID: 30587542 Free PMC article.
-
T-box transcription regulator Tbr2 is essential for the formation and maintenance of Opn4/melanopsin-expressing intrinsically photosensitive retinal ganglion cells.J Neurosci. 2014 Sep 24;34(39):13083-95. doi: 10.1523/JNEUROSCI.1027-14.2014. J Neurosci. 2014. PMID: 25253855 Free PMC article.
-
M1 ipRGCs Influence Visual Function through Retrograde Signaling in the Retina.J Neurosci. 2016 Jul 6;36(27):7184-97. doi: 10.1523/JNEUROSCI.3500-15.2016. J Neurosci. 2016. PMID: 27383593 Free PMC article.
-
Intrinsically photosensitive retinal ganglion cells: many subtypes, diverse functions.Trends Neurosci. 2011 Nov;34(11):572-80. doi: 10.1016/j.tins.2011.07.001. Epub 2011 Aug 3. Trends Neurosci. 2011. PMID: 21816493 Free PMC article. Review.
-
Intrinsically photosensitive retinal ganglion cells.Rev Physiol Biochem Pharmacol. 2012;162:59-90. doi: 10.1007/112_2011_4. Rev Physiol Biochem Pharmacol. 2012. PMID: 22160822 Review.
Cited by
-
The retina of the lab rat: focus on retinal ganglion cells and photoreceptors.Front Neuroanat. 2022 Sep 23;16:994890. doi: 10.3389/fnana.2022.994890. eCollection 2022. Front Neuroanat. 2022. PMID: 36213609 Free PMC article. Review.
-
Adult Mouse Retina Explants: From ex vivo to in vivo Model of Central Nervous System Injuries.Front Mol Neurosci. 2020 Nov 25;13:599948. doi: 10.3389/fnmol.2020.599948. eCollection 2020. Front Mol Neurosci. 2020. PMID: 33324161 Free PMC article.
-
Subtype-specific survival and regeneration of retinal ganglion cells in response to injury.Front Cell Dev Biol. 2022 Aug 12;10:956279. doi: 10.3389/fcell.2022.956279. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36035999 Free PMC article. Review.
-
A pupillary contrast response in mice and humans: Neural mechanisms and visual functions.Neuron. 2024 Jul 17;112(14):2404-2422.e9. doi: 10.1016/j.neuron.2024.04.012. Epub 2024 May 1. Neuron. 2024. PMID: 38697114 Free PMC article.
-
Pan-retinal ganglion cell markers in mice, rats, and rhesus macaques.Zool Res. 2023 Jan 18;44(1):226-248. doi: 10.24272/j.issn.2095-8137.2022.308. Zool Res. 2023. PMID: 36594396 Free PMC article. Review.
References
-
- Baver SB, Pickard GE, Sollars PJ, Pickard GE (2008) Two types of melanopsin retinal ganglion cell differentially innervate the hypothalamic suprachiasmatic nucleus and the olivary pretectal nucleus. Eur J Neurosci 27:1763–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases