

Cytokinin functions as an asymmetric and anti-gravitropic signal in lateral roots
- PMID: 31387989
- PMCID: PMC6684572
- DOI: 10.1038/s41467-019-11483-4
Cytokinin functions as an asymmetric and anti-gravitropic signal in lateral roots
Abstract
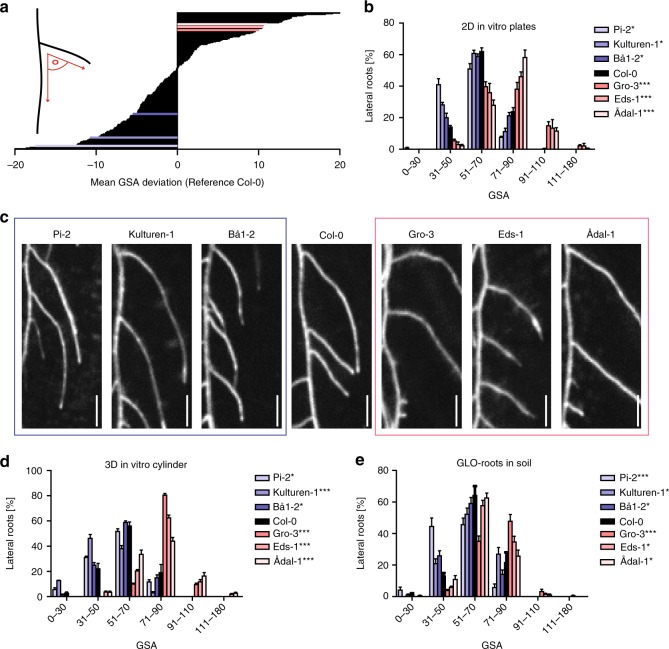
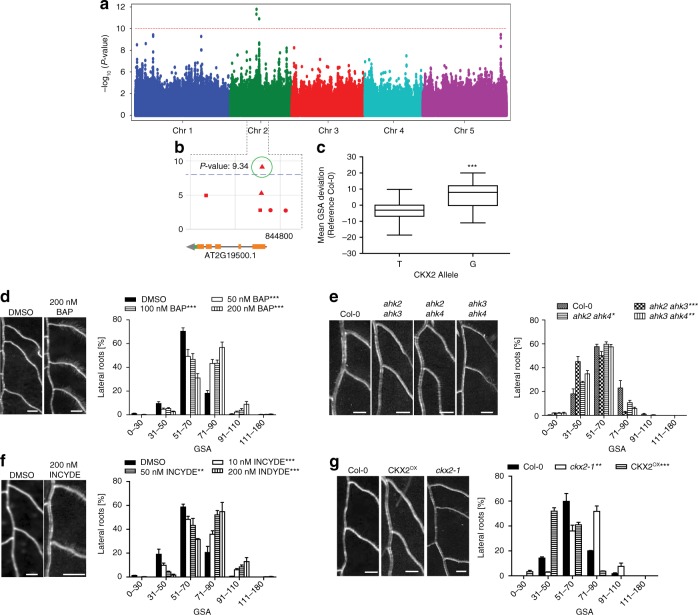
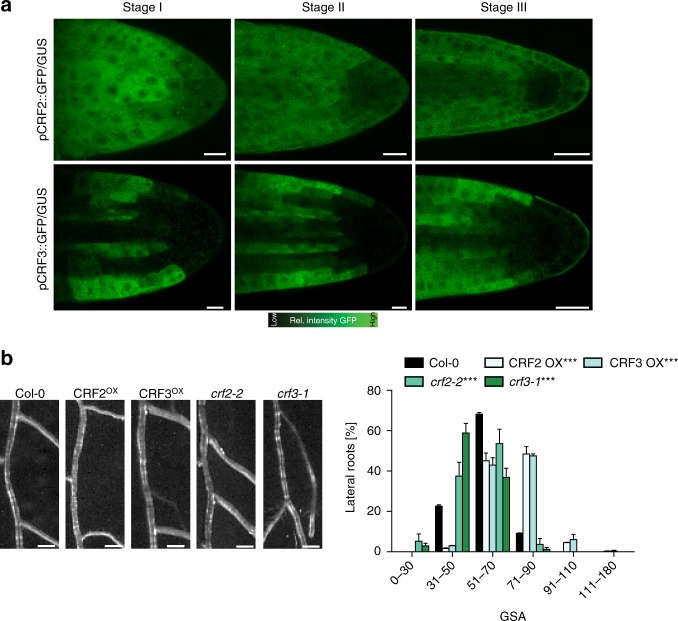
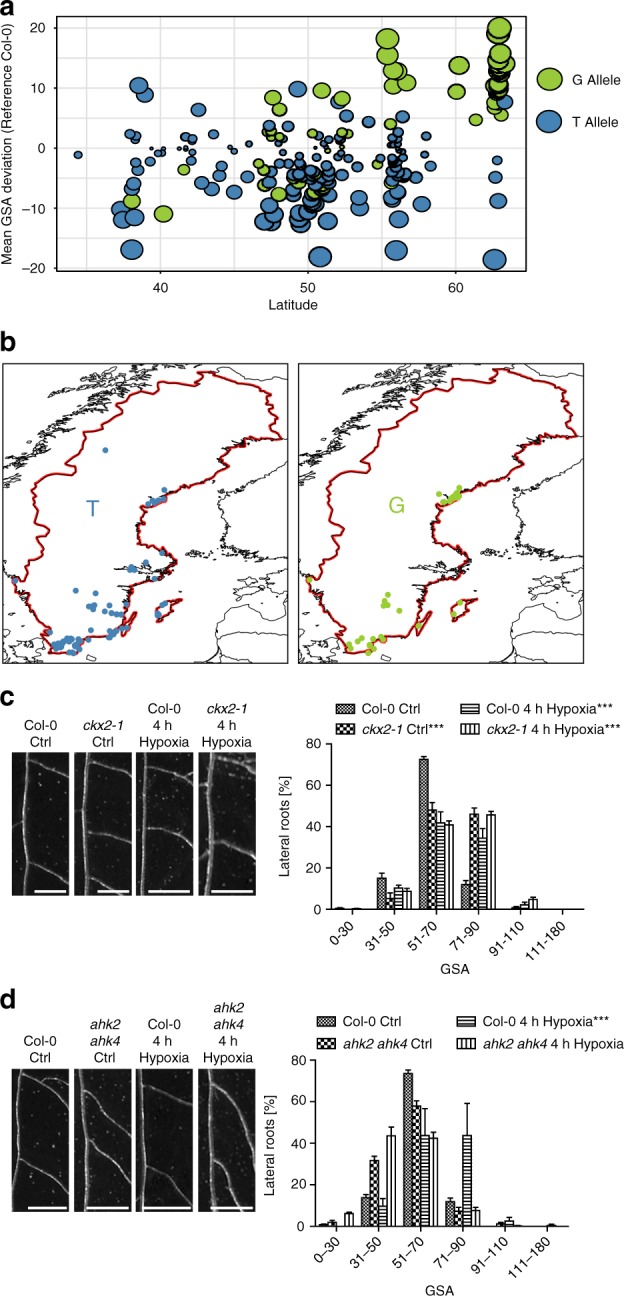
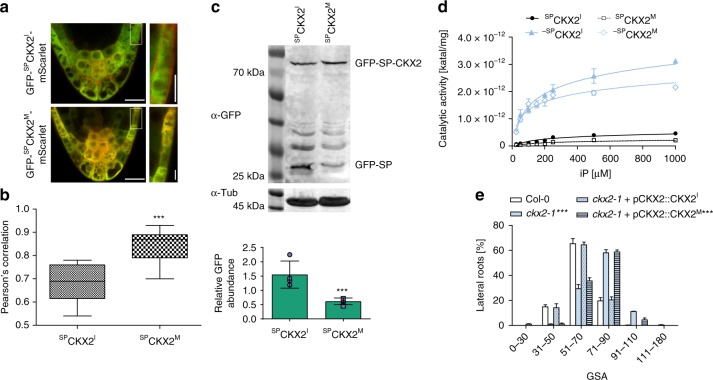
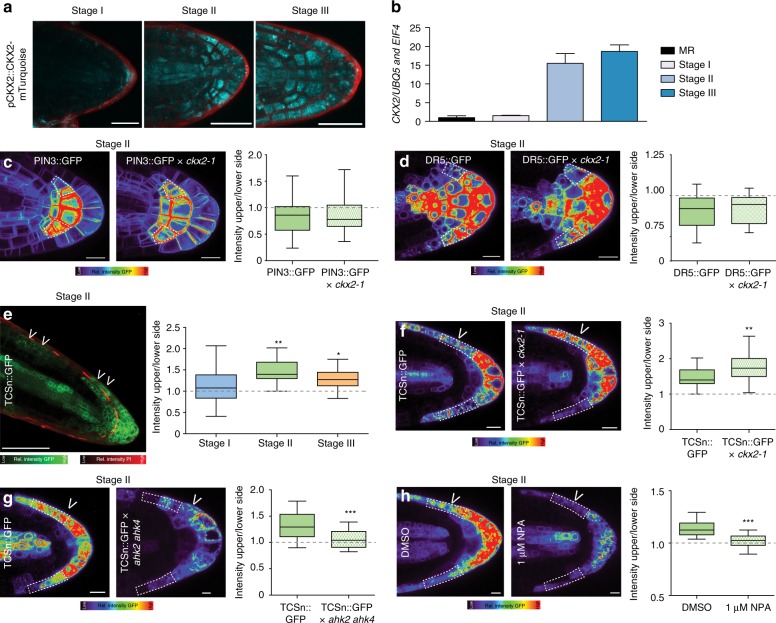
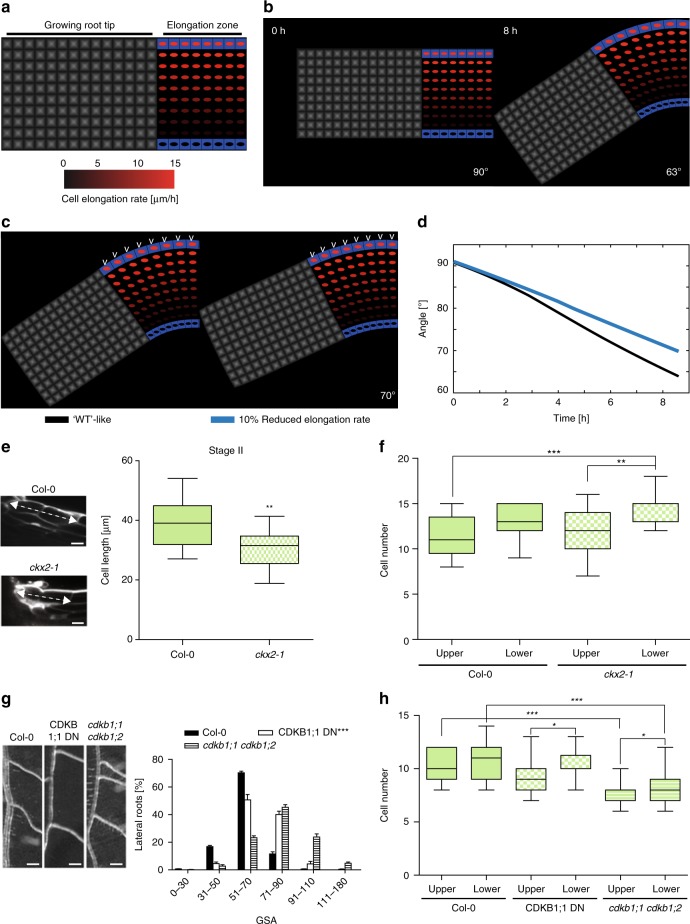
Directional organ growth allows the plant root system to strategically cover its surroundings. Intercellular auxin transport is aligned with the gravity vector in the primary root tips, facilitating downward organ bending at the lower root flank. Here we show that cytokinin signaling functions as a lateral root specific anti-gravitropic component, promoting the radial distribution of the root system. We performed a genome-wide association study and reveal that signal peptide processing of Cytokinin Oxidase 2 (CKX2) affects its enzymatic activity and, thereby, determines the degradation of cytokinins in natural Arabidopsis thaliana accessions. Cytokinin signaling interferes with growth at the upper lateral root flank and thereby prevents downward bending. Our interdisciplinary approach proposes that two phytohormonal cues at opposite organ flanks counterbalance each other's negative impact on growth, suppressing organ growth towards gravity and allow for radial expansion of the root system.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Adaptive Growth: Shaping Auxin-Mediated Root System Architecture.Trends Plant Sci. 2020 Feb;25(2):121-123. doi: 10.1016/j.tplants.2019.12.001. Epub 2019 Dec 13. Trends Plant Sci. 2020. PMID: 31843370
References
-
- Uga Y, et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Publ. Group. 2013;45:1097–1102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
