

Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber
- PMID: 31390991
- PMCID: PMC6686268
- DOI: 10.1186/s12870-019-1953-1
Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber
Abstract
Background: Aquaporin (AQP) proteins comprise a group of membrane intrinsic proteins (MIPs) that are responsible for transporting water and other small molecules, which is crucial for plant survival under stress conditions including salt stress. Despite the vital role of AQPs, little is known about them in cucumber (Cucumis sativus L.).
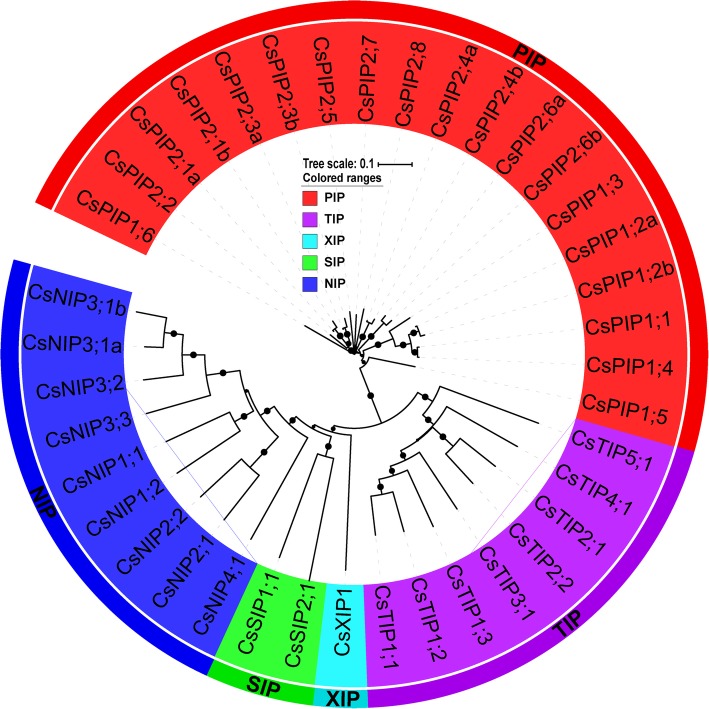
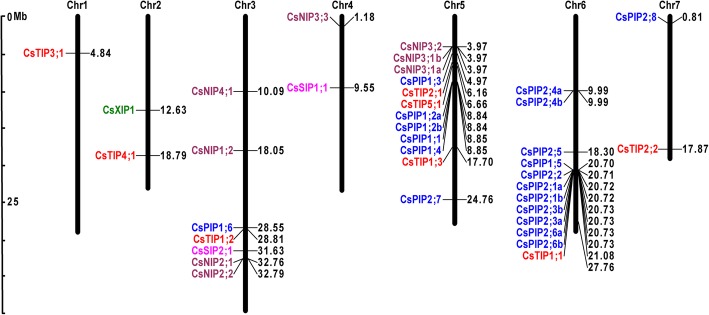
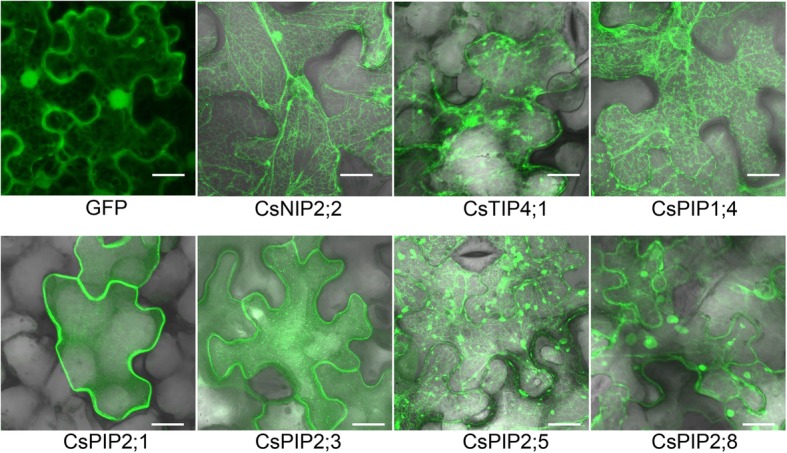
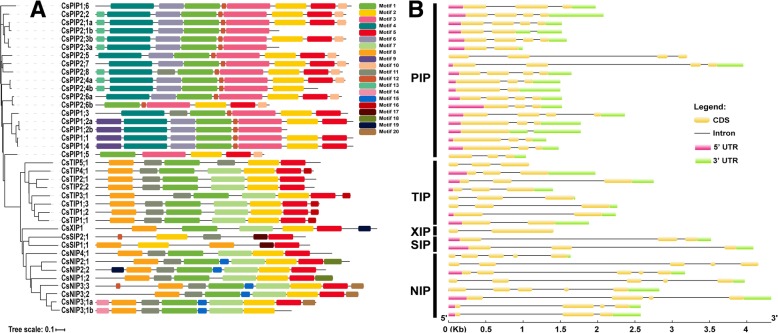
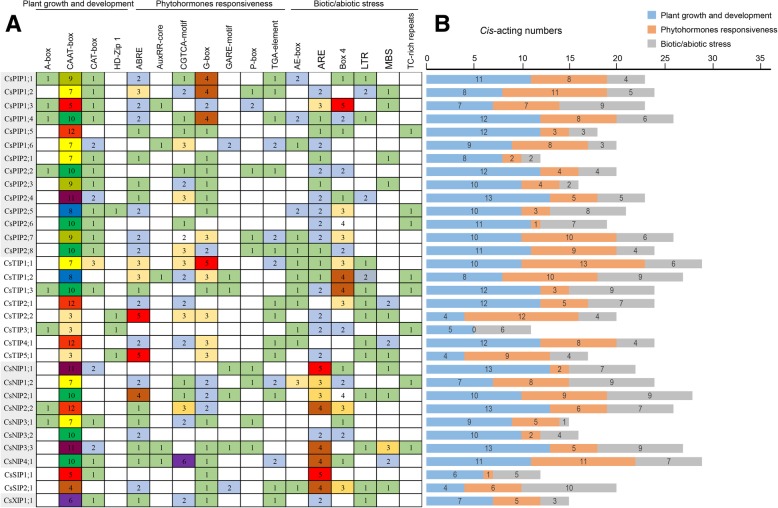
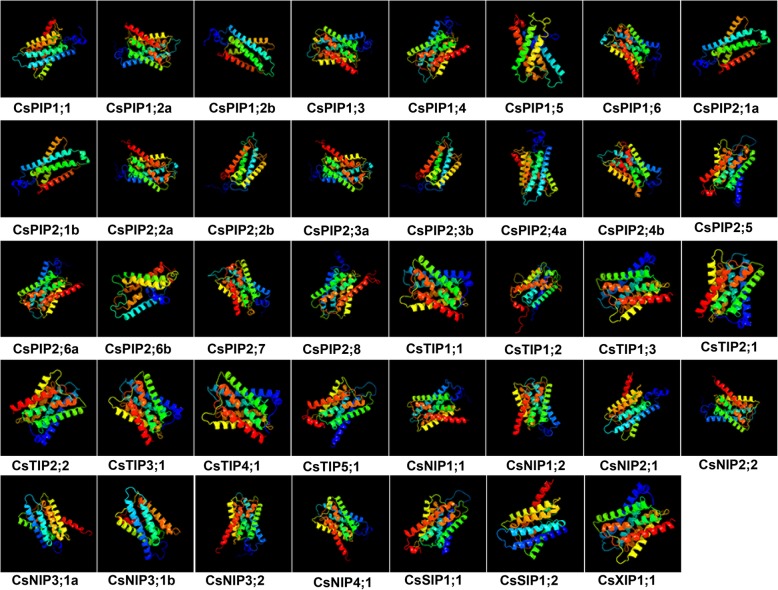
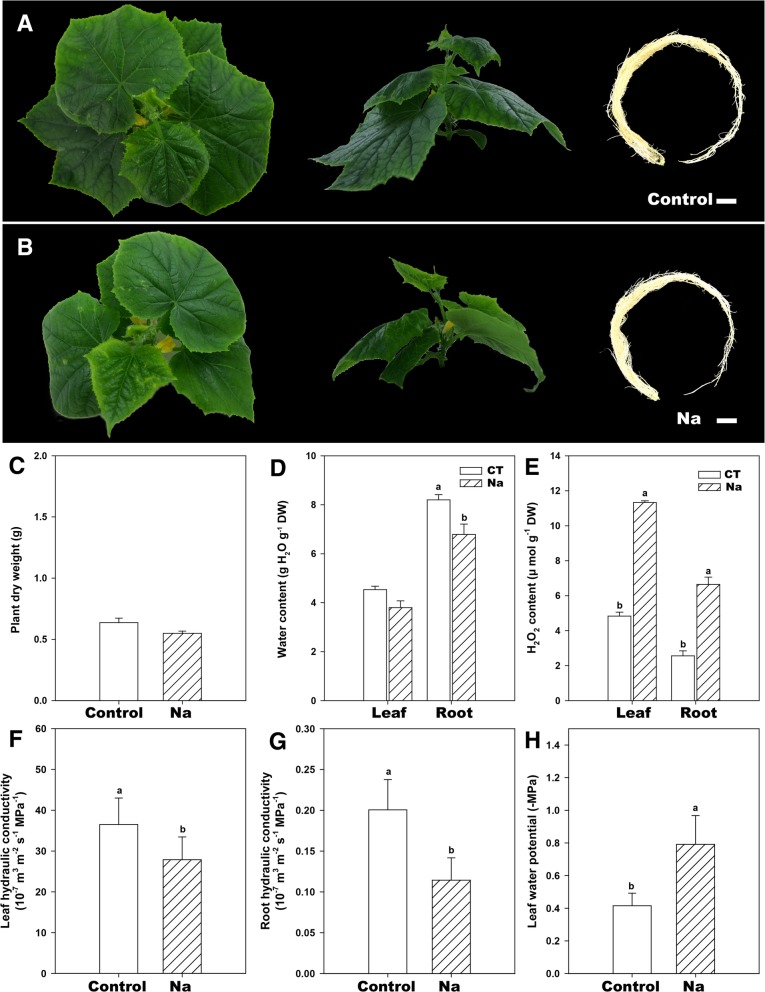
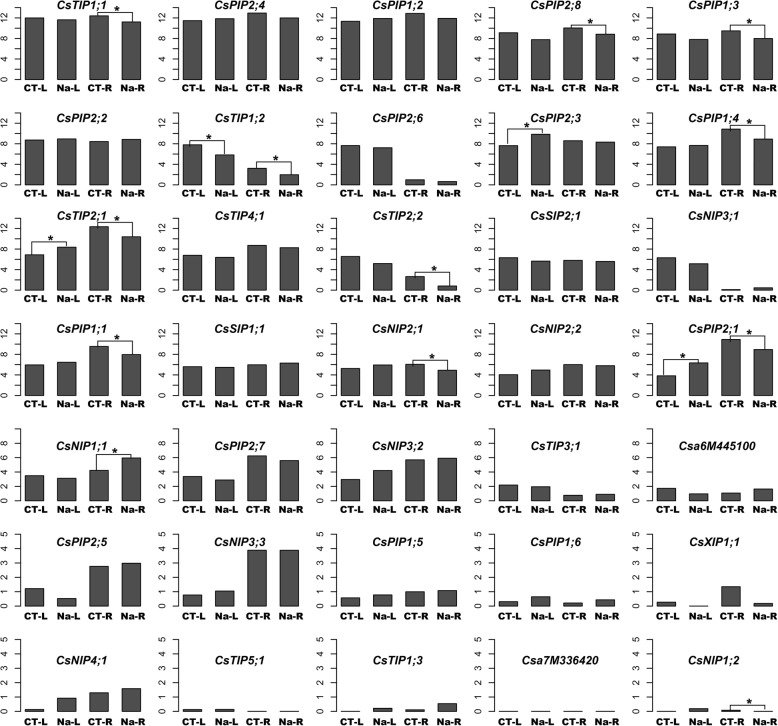
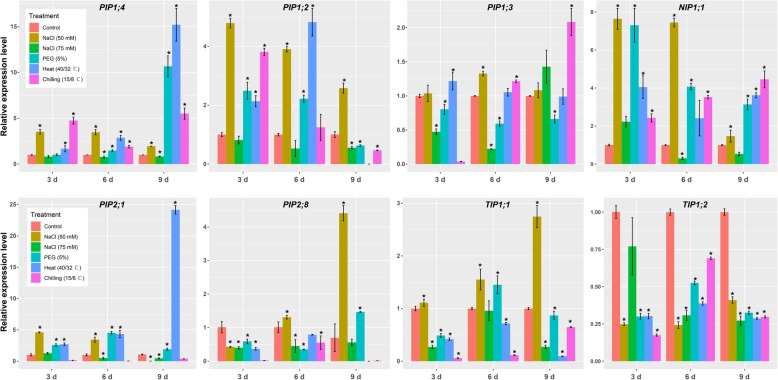
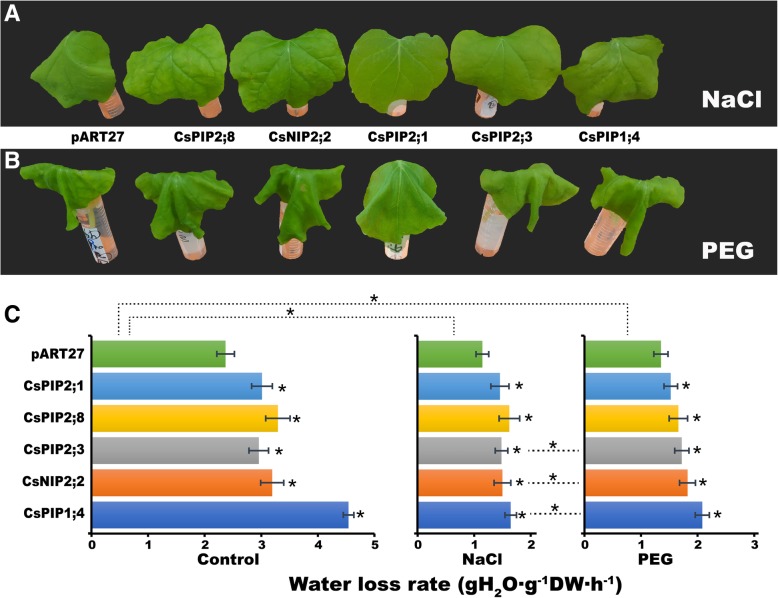
Results: In this study, we identified 39 aquaporin-encoding genes in cucumber that were separated by phylogenetic analysis into five sub-families (PIP, TIP, NIP, SIP, and XIP). Their substrate specificity was then assessed based on key amino acid residues such as the aromatic/Arginine (ar/R) selectivity filter, Froger's positions, and specificity-determining positions. The putative cis-regulatory motifs available in the promoter region of each AQP gene were analyzed and results revealed that their promoter regions contain many abiotic related cis-regulatory elements. Furthermore, analysis of previously released RNA-seq data revealed tissue- and treatment-specific expression patterns of cucumber AQP genes (CsAQPs). Three aquaporins (CsTIP1;1, CsPIP2;4, and CsPIP1;2) were the most transcript abundance genes, with CsTIP1;1 showing the highest expression levels among all aquaporins. Subcellular localization analysis in Nicotiana benthamiana epidermal cells revealed the diverse and broad array of sub-cellular localizations of CsAQPs. We then performed RNA-seq to identify the expression pattern of CsAQPs under salt stress and found a general decreased expression level of root CsAQPs. Moreover, qRT-PCR revealed rapid changes in the expression levels of CsAQPs in response to diverse abiotic stresses including salt, polyethylene glycol (PEG)-6000, heat, and chilling stresses. Additionally, transient expression of AQPs in N. benthamiana increased leaf water loss rate, suggesting their potential roles in the regulation of plant water status under stress conditions.
Conclusions: Our results indicated that CsAQPs play important roles in response to salt stress. The genome-wide identification and primary function characterization of cucumber aquaporins provides insight to elucidate the complexity of the AQP gene family and their biological functions in cucumber.
Keywords: Abiotic stresses; Gene structure; Phylogenetic analysis; Protein characterization; RNA-seq; Water loss rate.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Yin J, Jia J, Lian Z, Hu Y, Guo J, Huo H, Zhu Y, Gong H. Silicon enhances the salt tolerance of cucumber through increasing polyamine accumulation and decreasing oxidative damage. Ecotox Environ Safe. 2019;169:8–17. - PubMed
-
- Aroca R, Porcel R, Ruiz-Lozano JM. Regulation of root water uptake under abiotic stress conditions. J Exp Bot. 2011;63(1):43–57. - PubMed
-
- Johansson I, Karlsson M, Johanson U, Larsson C, Kjellbom P. The role of aquaporins in cellular and whole plant water balance. Biochim Biophys Acta. 2000;1465(1–2):324–342. - PubMed
-
- Fox AR, Maistriaux LC, Chaumont F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant Sci. 2017;264:179–187. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
