

Suppression of Mutant Protein Expression in SCA3 and SCA1 Mice Using a CAG Repeat-Targeting Antisense Oligonucleotide
- PMID: 31394429
- PMCID: PMC6695277
- DOI: 10.1016/j.omtn.2019.07.004
Suppression of Mutant Protein Expression in SCA3 and SCA1 Mice Using a CAG Repeat-Targeting Antisense Oligonucleotide
Abstract
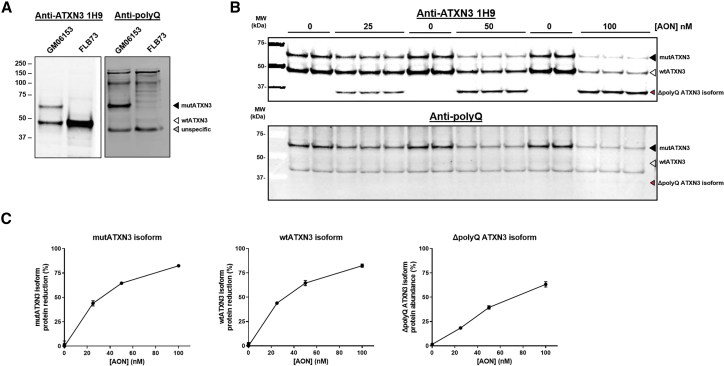
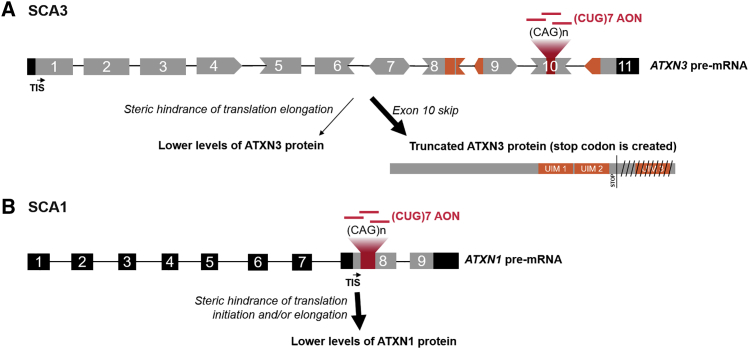
Spinocerebellar ataxia type 3 (SCA3) and type 1 (SCA1) are dominantly inherited neurodegenerative disorders that are currently incurable. Both diseases are caused by a CAG-repeat expansion in exon 10 of the Ataxin-3 and exon 8 of the Ataxin-1 gene, respectively, encoding an elongated polyglutamine tract that confers toxic properties to the resulting proteins. We have previously shown lowering of the pathogenic polyglutamine protein in Huntington's disease mouse models using (CUG)7, a CAG repeat-targeting antisense oligonucleotide. Here we evaluated the therapeutic capacity of (CUG)7 for SCA3 and SCA1, in vitro in patient-derived cell lines and in vivo in representative mouse models. Repeated intracerebroventricular (CUG)7 administration resulted in a significant reduction of mutant Ataxin-3 and Ataxin-1 proteins throughout the brain of SCA3 and SCA1 mouse models, respectively. Furthermore, in both a SCA3 patient cell line and the MJD84.2 mouse model, (CUG)7 induced formation of a truncated Ataxin-3 protein species lacking the polyglutamine stretch, likely arising from (CUG)7-mediated exon 10 skipping. In contrast, skipping of exon 8 of Ataxin-1 did not significantly contribute to the Ataxin-1 protein reduction observed in (CUG)7-treated SCA1154Q/2Q mice. These findings support the therapeutic potential of a single CAG repeat-targeting AON for the treatment of multiple polyglutamine disorders.
Keywords: CAG repeat; SCA1; SCA3; antisense oligonucleotide; exon skip; polyglutamine disorders.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Antisense Oligonucleotide-Mediated Removal of the Polyglutamine Repeat in Spinocerebellar Ataxia Type 3 Mice.Mol Ther Nucleic Acids. 2017 Sep 15;8:232-242. doi: 10.1016/j.omtn.2017.06.019. Epub 2017 Jun 29. Mol Ther Nucleic Acids. 2017. PMID: 28918024 Free PMC article.
-
Ataxin-3 protein modification as a treatment strategy for spinocerebellar ataxia type 3: removal of the CAG containing exon.Neurobiol Dis. 2013 Oct;58:49-56. doi: 10.1016/j.nbd.2013.04.019. Epub 2013 May 6. Neurobiol Dis. 2013. PMID: 23659897
-
Targeting several CAG expansion diseases by a single antisense oligonucleotide.PLoS One. 2011;6(9):e24308. doi: 10.1371/journal.pone.0024308. Epub 2011 Sep 1. PLoS One. 2011. PMID: 21909428 Free PMC article.
-
Progress in pathogenesis studies of spinocerebellar ataxia type 1.Philos Trans R Soc Lond B Biol Sci. 1999 Jun 29;354(1386):1079-81. doi: 10.1098/rstb.1999.0462. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10434309 Free PMC article. Review.
-
Clinical features, neurogenetics and neuropathology of the polyglutamine spinocerebellar ataxias type 1, 2, 3, 6 and 7.Prog Neurobiol. 2013 May;104:38-66. doi: 10.1016/j.pneurobio.2013.01.001. Epub 2013 Feb 21. Prog Neurobiol. 2013. PMID: 23438480 Review.
Cited by
-
Allele-specific targeting of mutant ataxin-3 by antisense oligonucleotides in SCA3-iPSC-derived neurons.Mol Ther Nucleic Acids. 2021 Nov 19;27:99-108. doi: 10.1016/j.omtn.2021.11.015. eCollection 2022 Mar 8. Mol Ther Nucleic Acids. 2021. PMID: 34938609 Free PMC article.
-
Accurate Quantification of Mutant and Wild-Type polyQ Proteins Using Simple Western Capillary Immunoassays.Mol Neurobiol. 2025 May 31. doi: 10.1007/s12035-025-05089-9. Online ahead of print. Mol Neurobiol. 2025. PMID: 40450087
-
Progress of macular atrophy during 30 months' follow-up in a patient with spinocerebellar ataxia type1 (SCA1).Doc Ophthalmol. 2021 Feb;142(1):87-98. doi: 10.1007/s10633-020-09782-z. Epub 2020 Jul 9. Doc Ophthalmol. 2021. PMID: 32648025
-
Construction of a trio-based structural variation panel utilizing activated T lymphocytes and long-read sequencing technology.Commun Biol. 2022 Sep 20;5(1):991. doi: 10.1038/s42003-022-03953-1. Commun Biol. 2022. PMID: 36127505 Free PMC article.
-
Antisense Oligonucleotide Therapeutic Approach for Suppression of Ataxin-1 Expression: A Safety Assessment.Mol Ther Nucleic Acids. 2020 Sep 4;21:1006-1016. doi: 10.1016/j.omtn.2020.07.030. Epub 2020 Jul 25. Mol Ther Nucleic Acids. 2020. PMID: 32818920 Free PMC article.
References
-
- Bird T.D. Hereditary Ataxia Overview. In: Adam M.P., Ardinger H.H., Pagon R.A., Wallace S.E., editors. GeneReviews. University of Washington,; Seattle: 1993. pp. 1993–2019. - PubMed
-
- van de Warrenburg B.P., Sinke R.J., Verschuuren-Bemelmans C.C., Scheffer H., Brunt E.R., Ippel P.F., Maat-Kievit J.A., Dooijes D., Notermans N.C., Lindhout D. Spinocerebellar ataxias in the Netherlands: prevalence and age at onset variance analysis. Neurology. 2002;58:702–708. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
