

Monocarboxylate transporters in cancer
- PMID: 31395464
- PMCID: PMC7056923
- DOI: 10.1016/j.molmet.2019.07.006
Monocarboxylate transporters in cancer
Abstract
Background: Tumors are highly plastic metabolic entities composed of cancer and host cells that can adopt different metabolic phenotypes. For energy production, cancer cells may use 4 main fuels that are shuttled in 5 different metabolic pathways. Glucose fuels glycolysis that can be coupled to the tricarboxylic acid (TCA) cycle and oxidative phosphorylation (OXPHOS) in oxidative cancer cells or to lactic fermentation in proliferating and in hypoxic cancer cells. Lipids fuel lipolysis, glutamine fuels glutaminolysis, and lactate fuels the oxidative pathway of lactate, all of which are coupled to the TCA cycle and OXPHOS for energy production. This review focuses on the latter metabolic pathway.
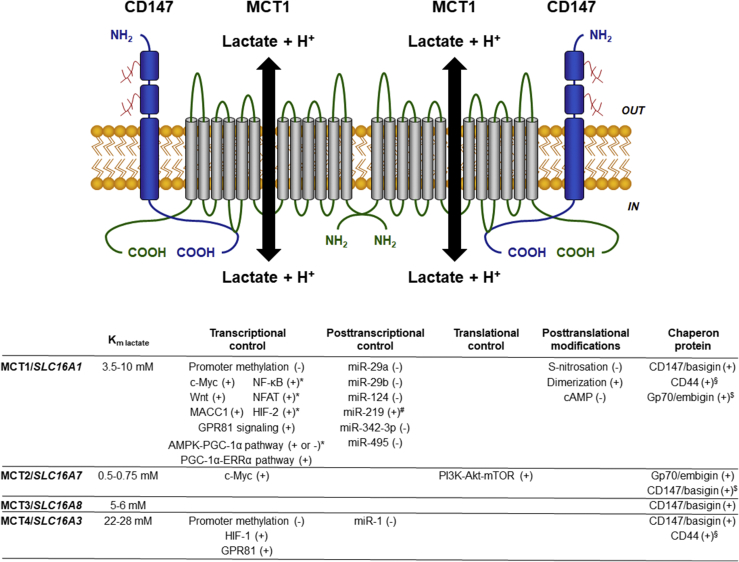
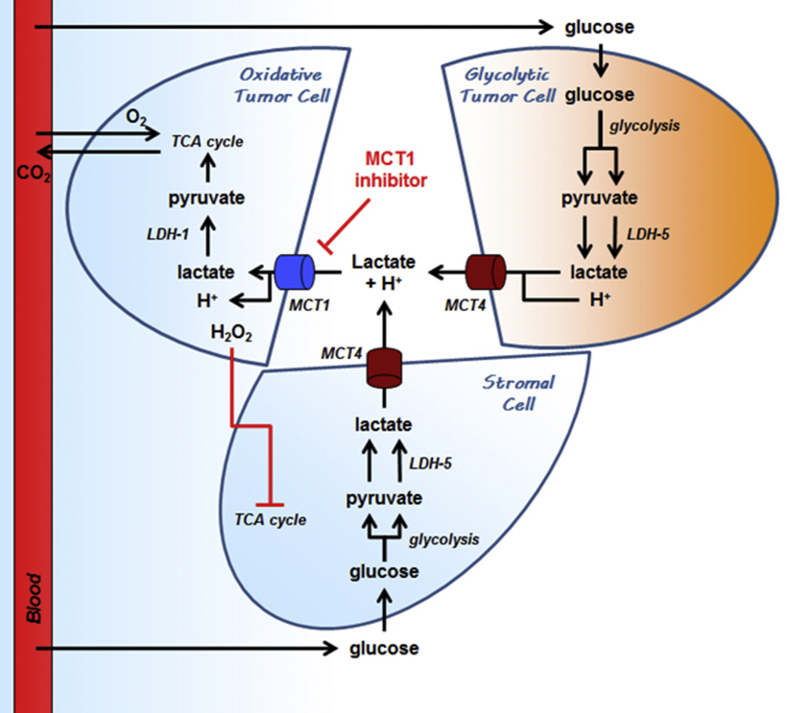
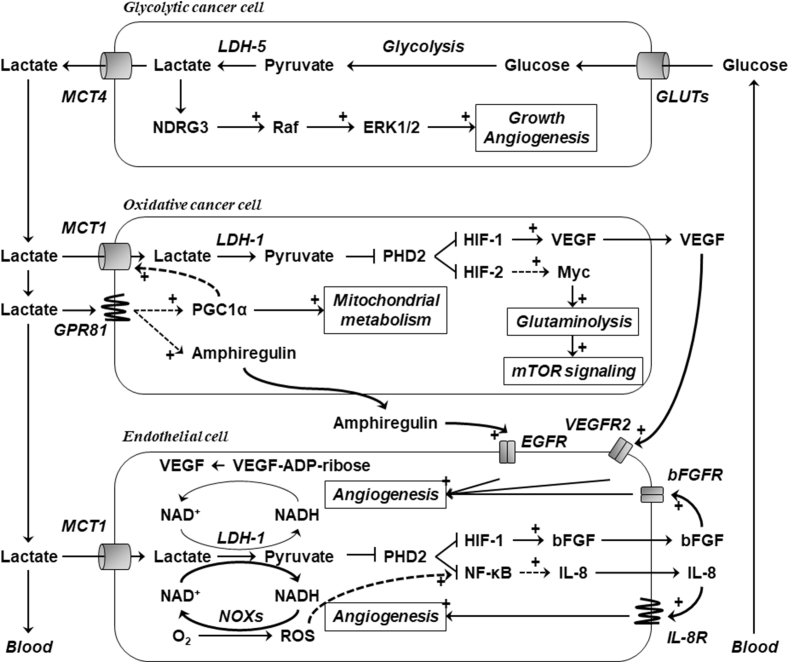
Scope of review: Lactate, which is prominently produced by glycolytic cells in tumors, was only recently recognized as a major fuel for oxidative cancer cells and as a signaling agent. Its exchanges across membranes are gated by monocarboxylate transporters MCT1-4. This review summarizes the current knowledge about MCT structure, regulation and functions in cancer, with a specific focus on lactate metabolism, lactate-induced angiogenesis and MCT-dependent cancer metastasis. It also describes lactate signaling via cell surface lactate receptor GPR81.
Major conclusions: Lactate and MCTs, especially MCT1 and MCT4, are important contributors to tumor aggressiveness. Analyses of MCT-deficient (MCT+/- and MCT-/-) animals and (MCT-mutated) humans indicate that they are druggable, with MCT1 inhibitors being in advanced development phase and MCT4 inhibitors still in the discovery phase. Imaging lactate fluxes non-invasively using a lactate tracer for positron emission tomography would further help to identify responders to the treatments.
Keywords: Angiogenesis; Cancer metabolism; GPR81; Metabolic symbiosis; Metastasis; Monocarboxylate transporters (MCTs).
Copyright © 2019 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Walenta S., Snyder S., Haroon Z.A., Braun R.D., Amin K., Brizel D. Tissue gradients of energy metabolites mirror oxygen tension gradients in a rat mammary carcinoma model. International Journal of Radiation Oncology, Biology, Physics. 2001;51:840–848. - PubMed
-
- Halestrap A.P., Wilson M.C. The monocarboxylate transporter family--role and regulation. IUBMB Life. 2012;64:109–119. - PubMed
-
- Halestrap A.P. The monocarboxylate transporter family--Structure and functional characterization. IUBMB Life. 2012;64:1–9. - PubMed
-
- Pinheiro C., Longatto-Filho A., Azevedo-Silva J., Casal M., Schmitt F.C., Baltazar F. Role of monocarboxylate transporters in human cancers: state of the art. Journal of Bioenergetics and Biomembranes. 2012;44:127–139. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
