

Rationale for targeting BCL6 in MLL-rearranged acute lymphoblastic leukemia
- PMID: 31395741
- PMCID: PMC6719625
- DOI: 10.1101/gad.327593.119
Rationale for targeting BCL6 in MLL-rearranged acute lymphoblastic leukemia
Abstract
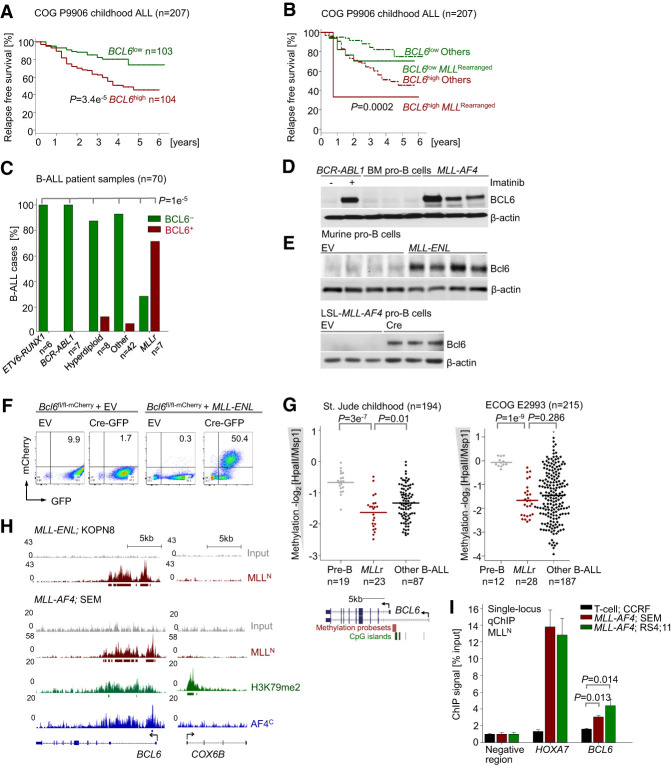
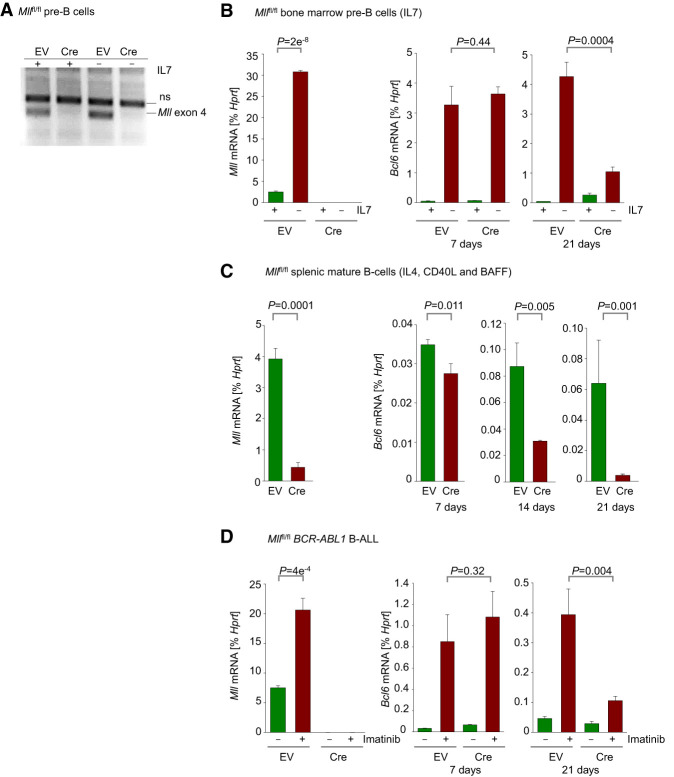
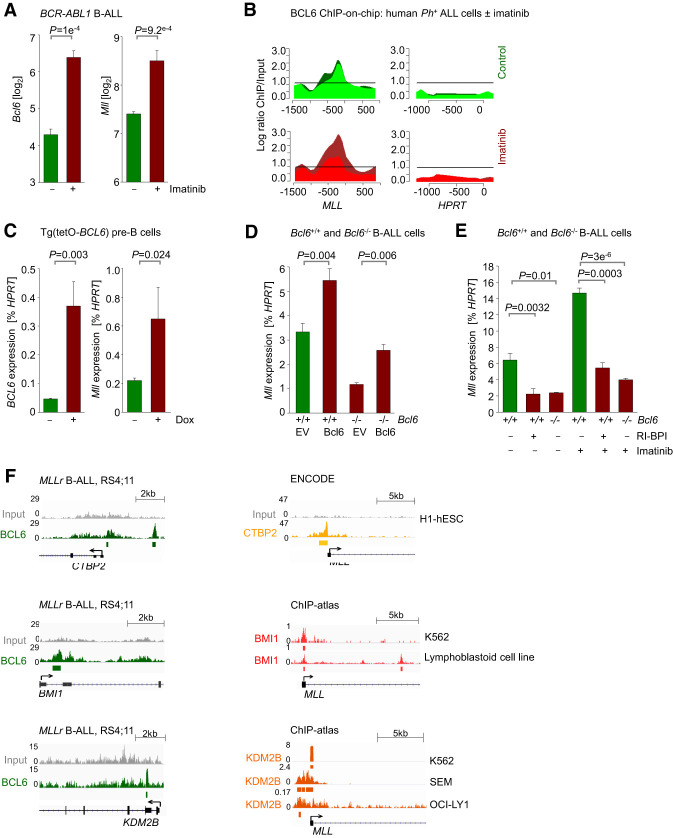
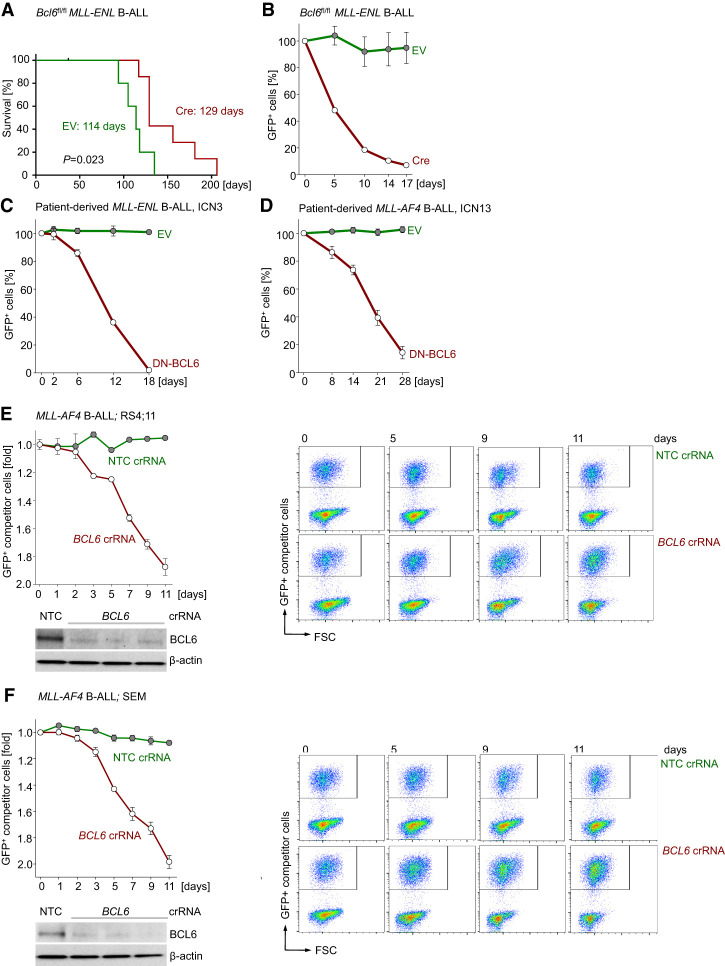
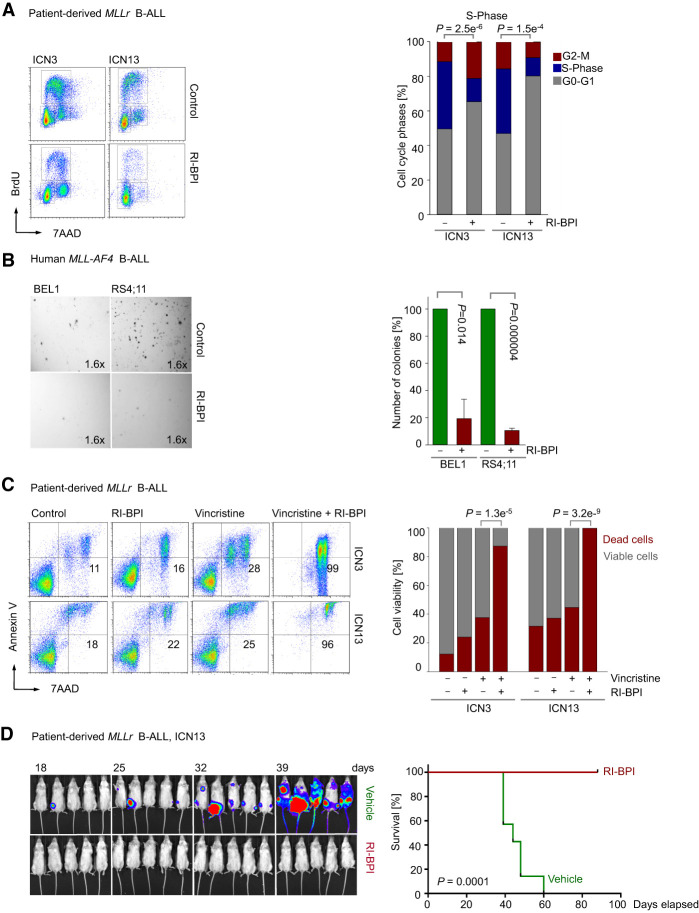
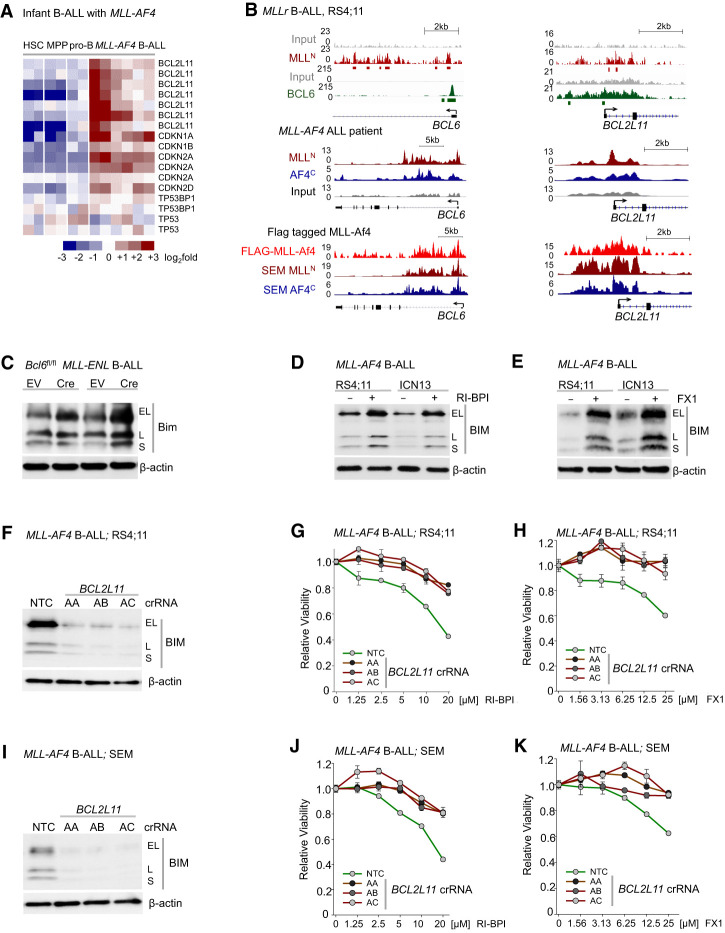
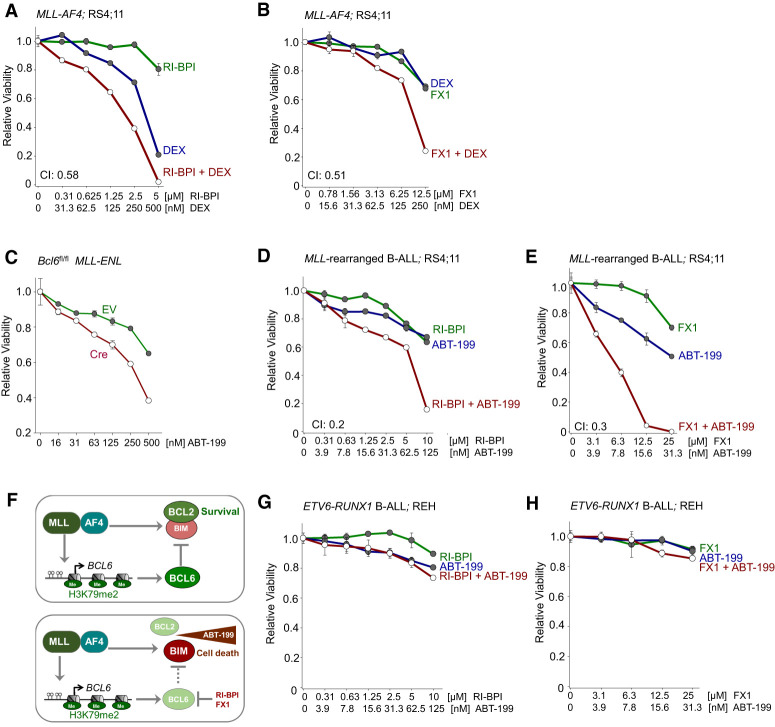
Chromosomal rearrangements of the mixed lineage leukemia (MLL) gene occur in ∼10% of B-cell acute lymphoblastic leukemia (B-ALL) and define a group of patients with dismal outcomes. Immunohistochemical staining of bone marrow biopsies from most of these patients revealed aberrant expression of BCL6, a transcription factor that promotes oncogenic B-cell transformation and drug resistance in B-ALL. Our genetic and ChIP-seq (chromatin immunoprecipitation [ChIP] combined with high-throughput sequencing) analyses showed that MLL-AF4 and MLL-ENL fusions directly bound to the BCL6 promoter and up-regulated BCL6 expression. While oncogenic MLL fusions strongly induced aberrant BCL6 expression in B-ALL cells, germline MLL was required to up-regulate Bcl6 in response to physiological stimuli during normal B-cell development. Inducible expression of Bcl6 increased MLL mRNA levels, which was reversed by genetic deletion and pharmacological inhibition of Bcl6, suggesting a positive feedback loop between MLL and BCL6. Highlighting the central role of BCL6 in MLL-rearranged B-ALL, conditional deletion and pharmacological inhibition of BCL6 compromised leukemogenesis in transplant recipient mice and restored sensitivity to vincristine chemotherapy in MLL-rearranged B-ALL patient samples. Oncogenic MLL fusions strongly induced transcriptional activation of the proapoptotic BH3-only molecule BIM, while BCL6 was required to curb MLL-induced expression of BIM. Notably, peptide (RI-BPI) and small molecule (FX1) BCL6 inhibitors derepressed BIM and synergized with the BH3-mimetic ABT-199 in eradicating MLL-rearranged B-ALL cells. These findings uncover MLL-dependent transcriptional activation of BCL6 as a previously unrecognized requirement of malignant transformation by oncogenic MLL fusions and identified BCL6 as a novel target for the treatment of MLL-rearranged B-ALL.
Keywords: B cells; BCL6; BIM; MLL.
© 2019 Hurtz et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Baron BW, Nucifora G, McCabe N, Espinosa R III, Le Beau MM, McKeithan TW. 1993. Identification of the gene associated with the recurring chromosomal translocations t(3;14)(q27;q32) and t(3;22)(q27;q11) in B-cell lymphomas. Proc Natl Acad Sci 90: 5262–5266. 10.1073/pnas.90.11.5262 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA198089/CA/NCI NIH HHS/United States
- U54 CA231637/CA/NCI NIH HHS/United States
- R35 CA220499/CA/NCI NIH HHS/United States
- R01 CA236399/CA/NCI NIH HHS/United States
- MC_UU_00016/6/MRC_/Medical Research Council/United Kingdom
- R01 CA213138/CA/NCI NIH HHS/United States
- R01 CA214965/CA/NCI NIH HHS/United States
- R01 CA172558/CA/NCI NIH HHS/United States
- HHMI-55108547/HHMI/Howard Hughes Medical Institute/United States
- R37 CA233691/CA/NCI NIH HHS/United States
- R01 CA243386/CA/NCI NIH HHS/United States
- R01 CA204639/CA/NCI NIH HHS/United States
- R00 CA197498/CA/NCI NIH HHS/United States
- P01 CA066996/CA/NCI NIH HHS/United States
- R01 CA157644/CA/NCI NIH HHS/United States
- R01 CA178454/CA/NCI NIH HHS/United States
- R01 CA211614/CA/NCI NIH HHS/United States
- R35 CA197628/CA/NCI NIH HHS/United States
- R01 CA176745/CA/NCI NIH HHS/United States
- MC_UU_12009/6/MRC_/Medical Research Council/United Kingdom
- U10 CA180827/CA/NCI NIH HHS/United States
- R01 CA236626/CA/NCI NIH HHS/United States
- R01 CA137060/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases