

Knock-in of Mutated hTAU Causes Insulin Resistance, Inflammation and Proteostasis Disturbance in a Mouse Model of Frontotemporal Dementia
- PMID: 31396860
- PMCID: PMC6968995
- DOI: 10.1007/s12035-019-01722-6
Knock-in of Mutated hTAU Causes Insulin Resistance, Inflammation and Proteostasis Disturbance in a Mouse Model of Frontotemporal Dementia
Abstract
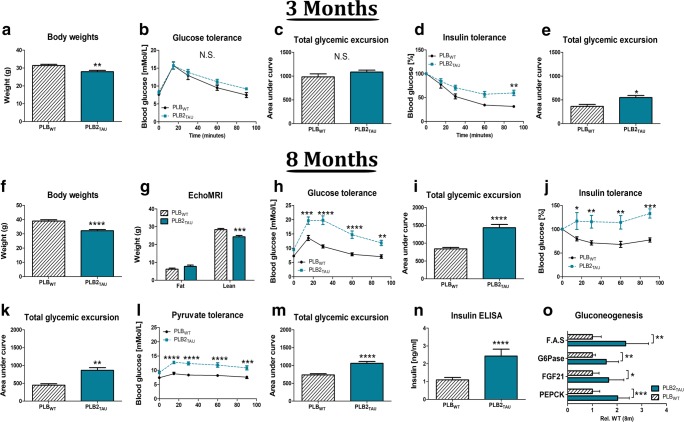
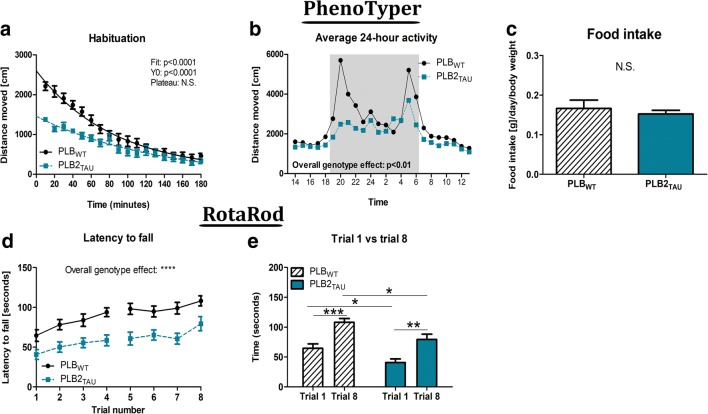
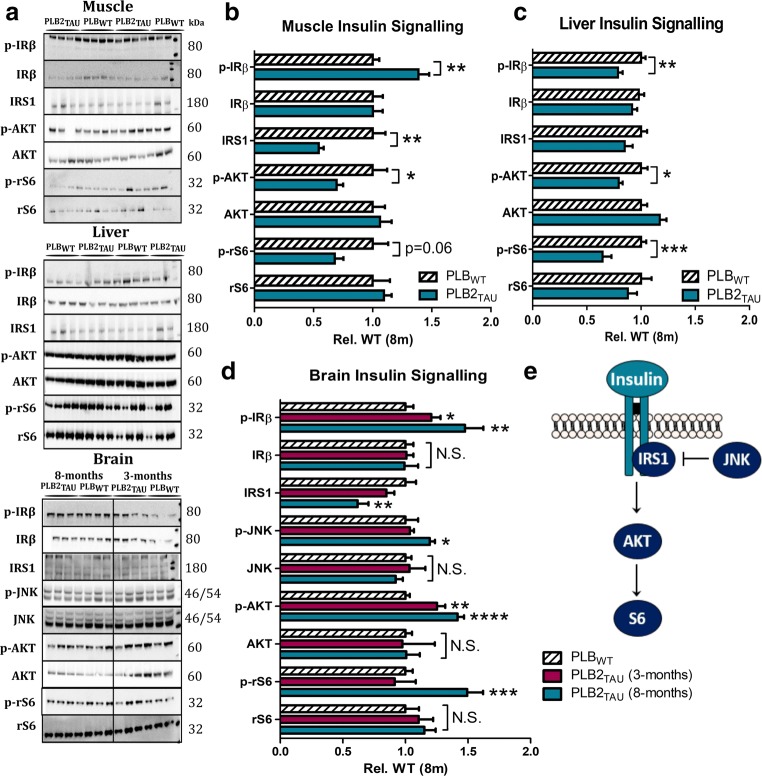
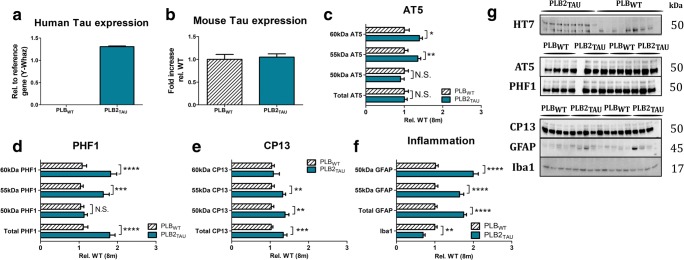
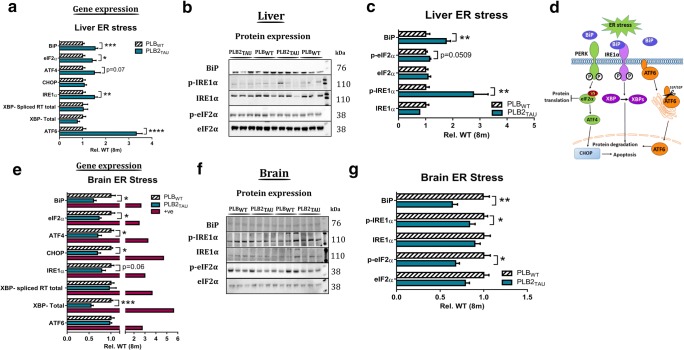
Diabetes and obesity have been implicated as risk factors for dementia. However, metabolic mechanisms and associated signalling pathways have not been investigated in detail in frontotemporal dementia. We therefore here characterised physiological, behavioural and molecular phenotypes of 3- and 8-month-old male tau knock-in (PLB2TAU) vs wild-type (PLBWT) mice. Homecage analysis suggested intact habituation but a dramatic reduction in exploratory activity in PLB2TAU mice. Deficits in motor strength were also observed. At 3 months, PLB2TAU mice displayed normal glucose handling but developed hyperglycaemia at 8 months, suggesting a progressive diabetic phenotype. Brain, liver and muscle tissue analyses confirmed tissue-specific deregulation of metabolic and homeostatic pathways. In brain, increased levels of phosphorylated tau and inflammation were detected alongside reduced ER regulatory markers, overall suggesting a downregulation in essential cellular defence pathways. We suggest that subtle neuronal expression of mutated human tau is sufficient to disturb systems metabolism and protein handling. Whether respective dysfunctions in tauopathy patients are also a consequence of tau pathology remains to be confirmed, but could offer new avenues for therapeutic interventions.
Keywords: Diabetes; ER stress; Glucose; Insulin; Knock-in; Proteinopathy; Transgenic; UPR.
Figures

References
-
- Yarchoan M, Toledo JB, Lee EB, Arvanitakis Z, Kazi H, Han LY, Louneva N, Lee VMY, Kim SF, Trojanowski JQ, Arnold SE. Abnormal serine phosphorylation of insulin receptor substrate 1 is associated with tau pathology in Alzheimer’s disease and tauopathies. Acta Neuropathol. 2014;128:679–689. doi: 10.1007/s00401-014-1328-5. - DOI - PMC - PubMed
-
- Marciniak E, Leboucher A, Caron E, Ahmed T, Tailleux A, Dumont J, Issad T, Gerhardt E, Pagesy P, Vileno M, Bournonville C, Hamdane M, Bantubungi K, Lancel S, Demeyer D, Eddarkaoui S, Vallez E, Vieau D, Humez S, Faivre E, Grenier-Boley B, Outeiro TF, Staels B, Amouyel P, Balschun D, Buee L, Blum D. Tau deletion promotes brain insulin resistance. J Exp Med. 2017;214:2257–2269. doi: 10.1084/jem.20161731. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
