

Prebiotics: Mechanisms and Preventive Effects in Allergy
- PMID: 31398959
- PMCID: PMC6722770
- DOI: 10.3390/nu11081841
Prebiotics: Mechanisms and Preventive Effects in Allergy
Abstract
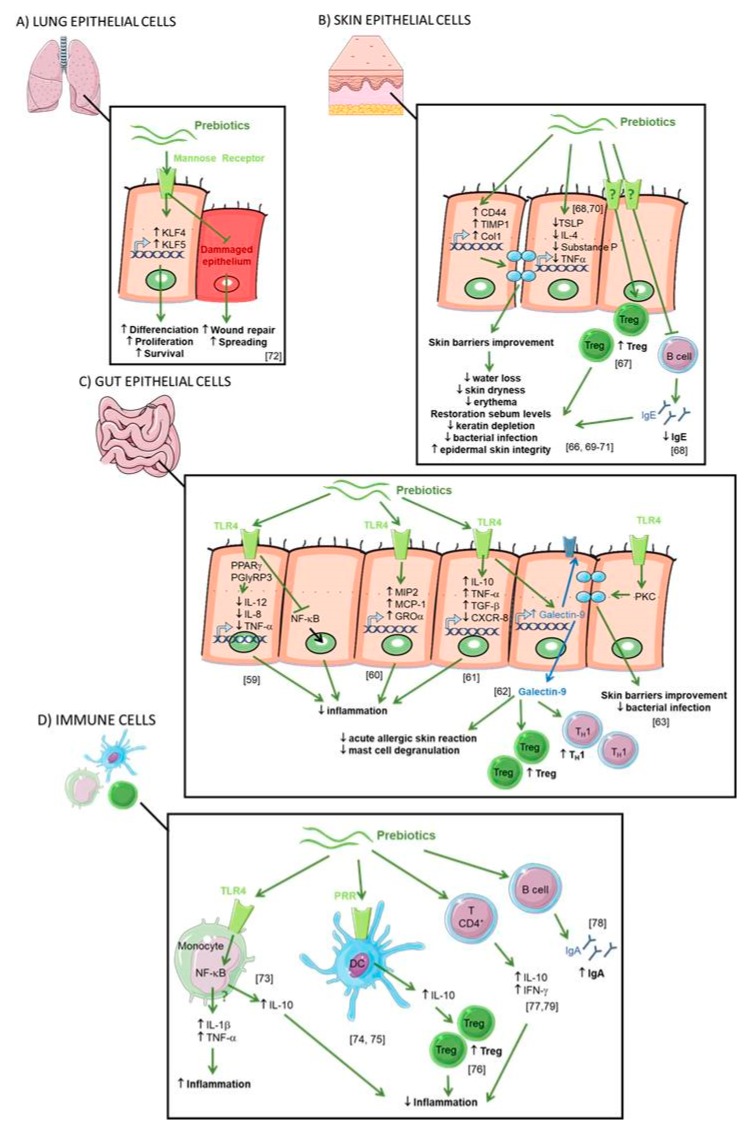
Allergic diseases now affect over 30% of individuals in many communities, particularly young children, underscoring the need for effective prevention strategies in early life. These allergic conditions have been linked to environmental and lifestyle changes driving the dysfunction of three interdependent biological systems: microbiota, epithelial barrier and immune system. While this is multifactorial, dietary changes are of particular interest in the altered establishment and maturation of the microbiome, including the associated profile of metabolites that modulate immune development and barrier function. Prebiotics are non-digestible food ingredients that beneficially influence the health of the host by 1) acting as a fermentable substrate for some specific commensal host bacteria leading to the release of short-chain fatty acids in the gut intestinal tract influencing many molecular and cellular processes; 2) acting directly on several compartments and specifically on different patterns of cells (epithelial and immune cells). Nutrients with prebiotic properties are therefore of central interest in allergy prevention for their potential to promote a more tolerogenic environment through these multiple pathways. Both observational studies and experimental models lend further credence to this hypothesis. In this review, we describe both the mechanisms and the therapeutic evidence from preclinical and clinical studies exploring the role of prebiotics in allergy prevention.
Keywords: allergy; clinical studies; epithelial barrier; immune system; mechanisms; microbiota; prebiotics; preclinical studies.
Conflict of interest statement
A.S., C.B. and S.B. declare no conflict of interest. M.B. reports the following: Scientific Advisory Board from the French Society of Nutrition, Paris, France. Research grants and speaker’s fees from Biostime Institute Nutrition and Care (BINC), Geneva, Switzerland; from the French Society of Allergy, Paris, France; from the French Society of Nutrition, Paris, France; from the French National Research Agency, Paris, France; from the Research, Education and Innovation program Food for Tomorrow/Cap Aliment, the Pays de la Loire Region, France; from interregional clinical research hospital program, France. S.L.P. reports the following: Scientific Advisory Board and speaker’s fees from Danone Nutricia, Utrecht, Netherlands and Biostime Institute Nutrition and Care (BINC), Geneva, Switzerland; consultancy fees from Bayer Dietary Supplements Division, Whippany, NJ, USA; speakers fees from Health World Inc., Queensland, Australia; research grants from the National Health and Medical Research Council (NHMRC) of Australia. D.J.P. reports the following: research grants from the National Health and Medical Research Council (NHMRC) of Australia, Foundation for Children, Australian Food Allergy Foundation and Telethon Perth Children’s Hospital Research Fund; and speaker’s fees from Abbott, Danone Nutricia and Nestle Nutrition Institute.
Figures
References
-
- Pawankar R., Canonica G.W., Holgate S.T., Lockey R. World Allergy Organization (WAO) white book on allergy. World Allergy Organization; Milwaukee, WI, USA: 2011.
-
- Werfel T., Allam J.P., Biedermann T., Eyerich K., Gilles S., Guttman-Yassky E., Hoetzenecker W., Knol E., Simon H.U., Wollenberg A., et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016;138:336–349. doi: 10.1016/j.jaci.2016.06.010. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
