

The Histone Deacetylase SIRT6 Restrains Transcription Elongation via Promoter-Proximal Pausing
- PMID: 31399344
- PMCID: PMC6907403
- DOI: 10.1016/j.molcel.2019.06.034
The Histone Deacetylase SIRT6 Restrains Transcription Elongation via Promoter-Proximal Pausing
Abstract
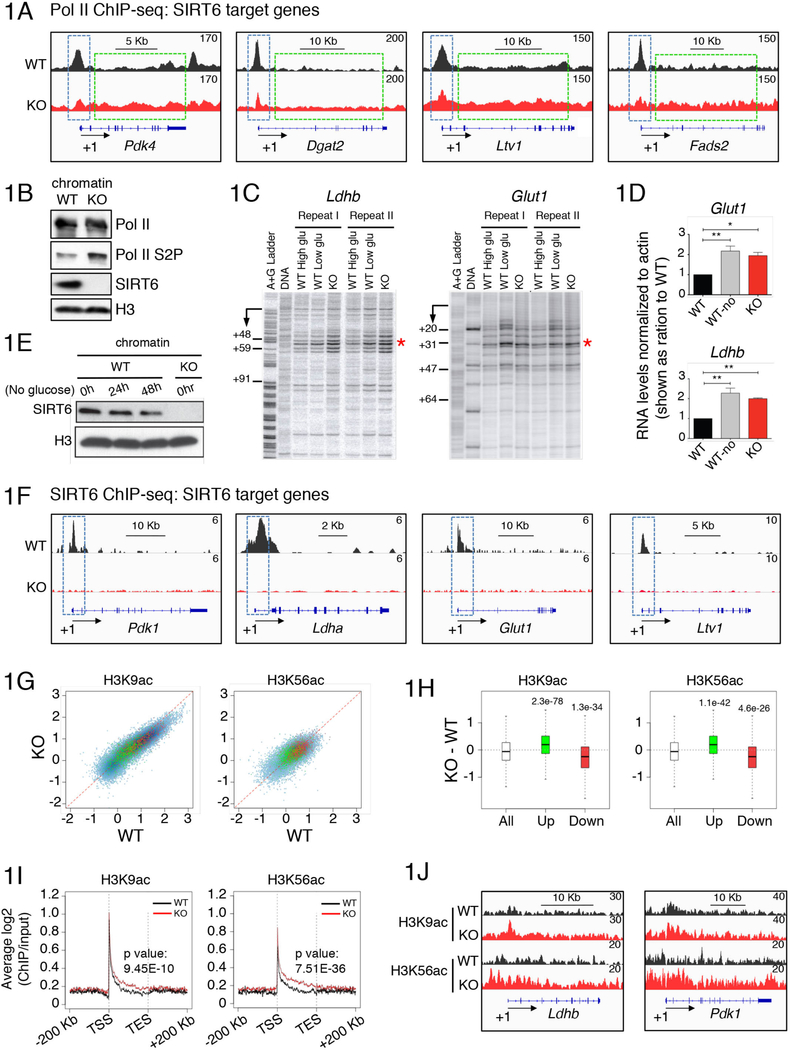
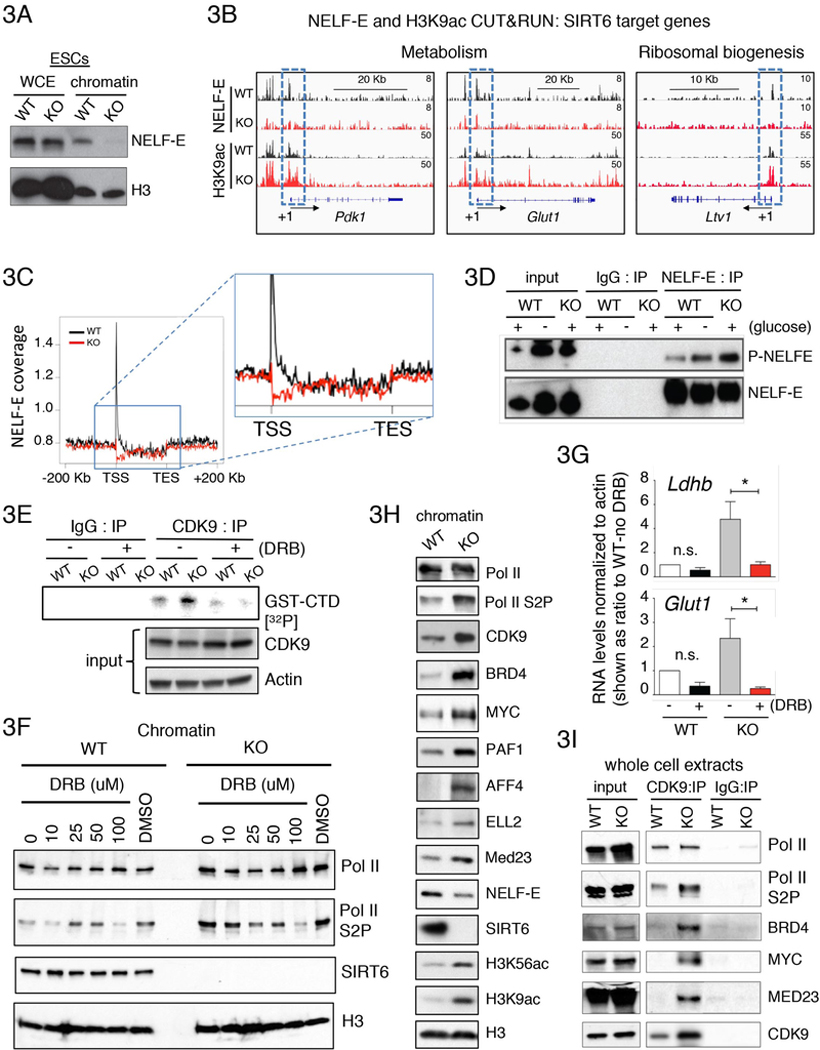
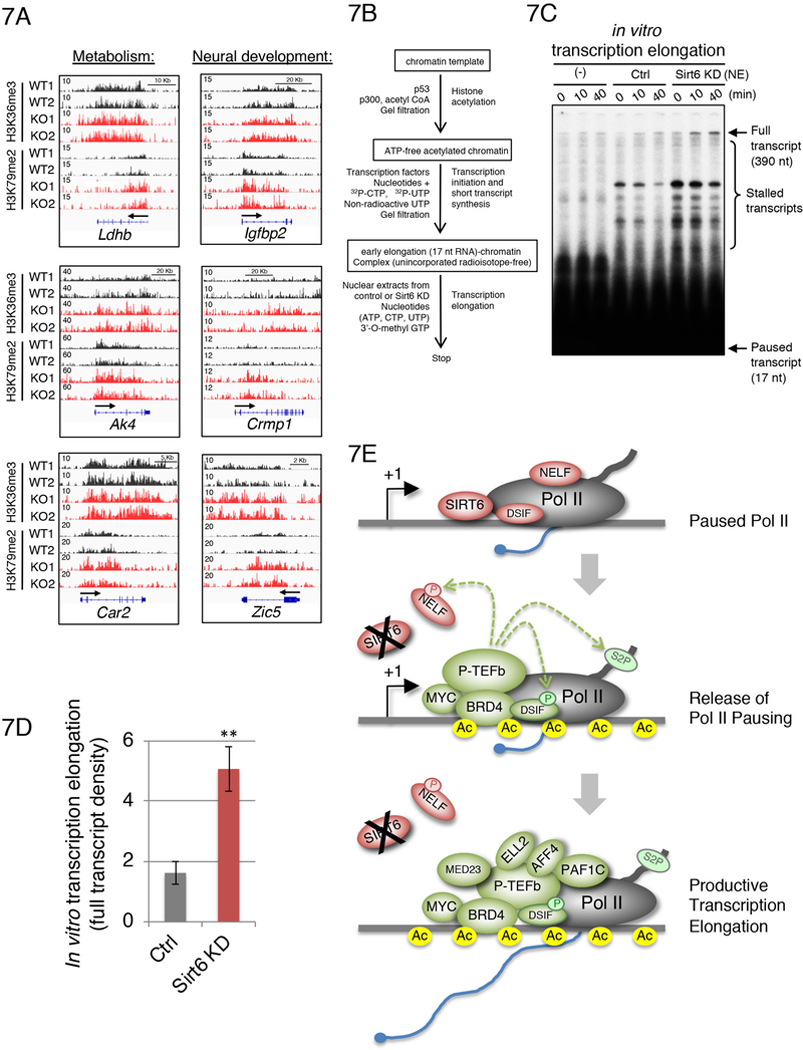
Transcriptional regulation in eukaryotes occurs at promoter-proximal regions wherein transcriptionally engaged RNA polymerase II (Pol II) pauses before proceeding toward productive elongation. The role of chromatin in pausing remains poorly understood. Here, we demonstrate that the histone deacetylase SIRT6 binds to Pol II and prevents the release of the negative elongation factor (NELF), thus stabilizing Pol II promoter-proximal pausing. Genetic depletion of SIRT6 or its chromatin deficiency upon glucose deprivation causes intragenic enrichment of acetylated histone H3 at lysines 9 (H3K9ac) and 56 (H3K56ac), activation of cyclin-dependent kinase 9 (CDK9)-that phosphorylates NELF and the carboxyl terminal domain of Pol II-and enrichment of the positive transcription elongation factors MYC, BRD4, PAF1, and the super elongation factors AFF4 and ELL2. These events lead to increased expression of genes involved in metabolism, protein synthesis, and embryonic development. Our results identified SIRT6 as a Pol II promoter-proximal pausing-dedicated histone deacetylase.
Keywords: BRD4; NELF; SIRT6; chromatin; epigenetics; histone deacetylation; transcription elongation; transcriptional pausing.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures




References
-
- Alland L, Muhle R, Hou H Jr., Potes J, Chin L, Schreiber-Agus N, and DePinho RA (1997). Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature 387, 49–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
