

Diurnal rhythms in gene expression in the prefrontal cortex in schizophrenia
- PMID: 31399567
- PMCID: PMC6689017
- DOI: 10.1038/s41467-019-11335-1
Diurnal rhythms in gene expression in the prefrontal cortex in schizophrenia
Abstract
Schizophrenia is associated with disrupted cognitive control and sleep-wake cycles. Here we identify diurnal rhythms in gene expression in the human dorsolateral prefrontal cortex (dlPFC), in schizophrenia and control subjects. We find significant diurnal (24 h) rhythms in control subjects, however, most of these transcripts are not rhythmic in subjects with schizophrenia. Instead, subjects with schizophrenia have a different set of rhythmic transcripts. The top pathways identified in transcripts rhythmic only in subjects with schizophrenia are associated with mitochondrial function. Importantly, these rhythms drive differential expression patterns of these and several other genes that have long been implicated in schizophrenia (including BDNF and GABAergic-related transcripts). Indeed, differential expression of these transcripts is only seen in subjects that died during the night, with no change in subjects that died during the day. These data provide insights into a potential mechanism that underlies changes in gene expression in the dlPFC with schizophrenia.
Conflict of interest statement
The authors declare no competing interests.
Figures
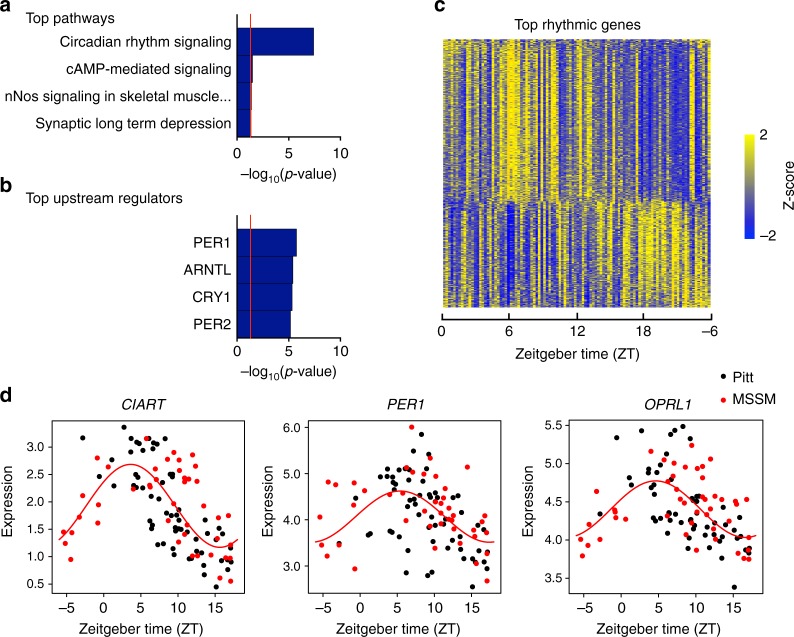
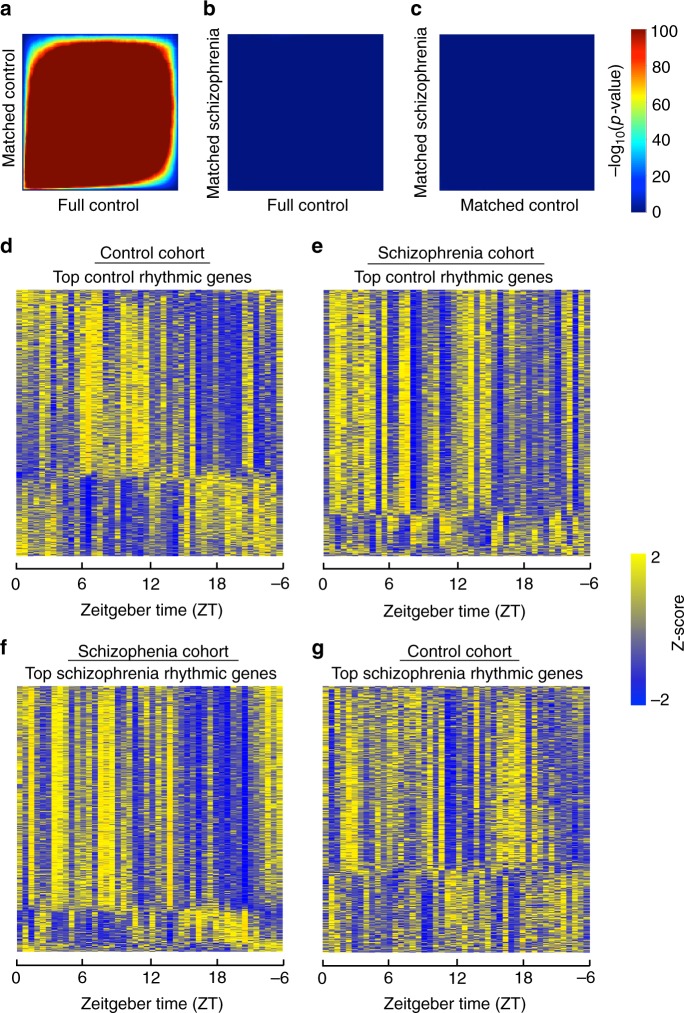
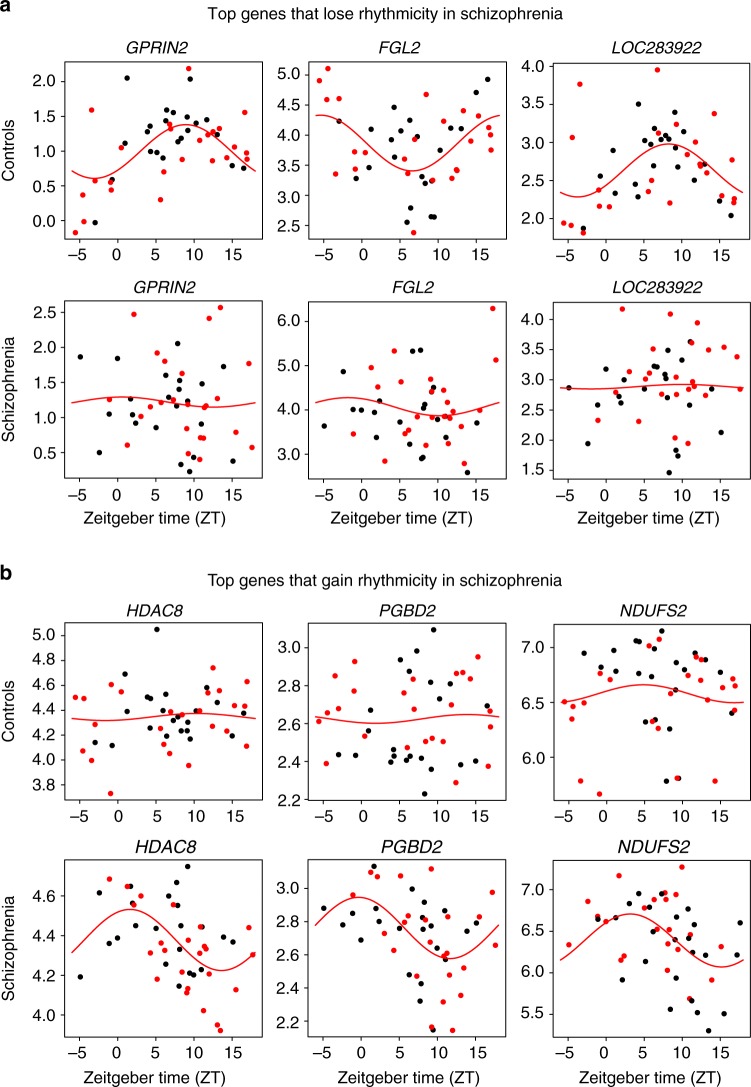
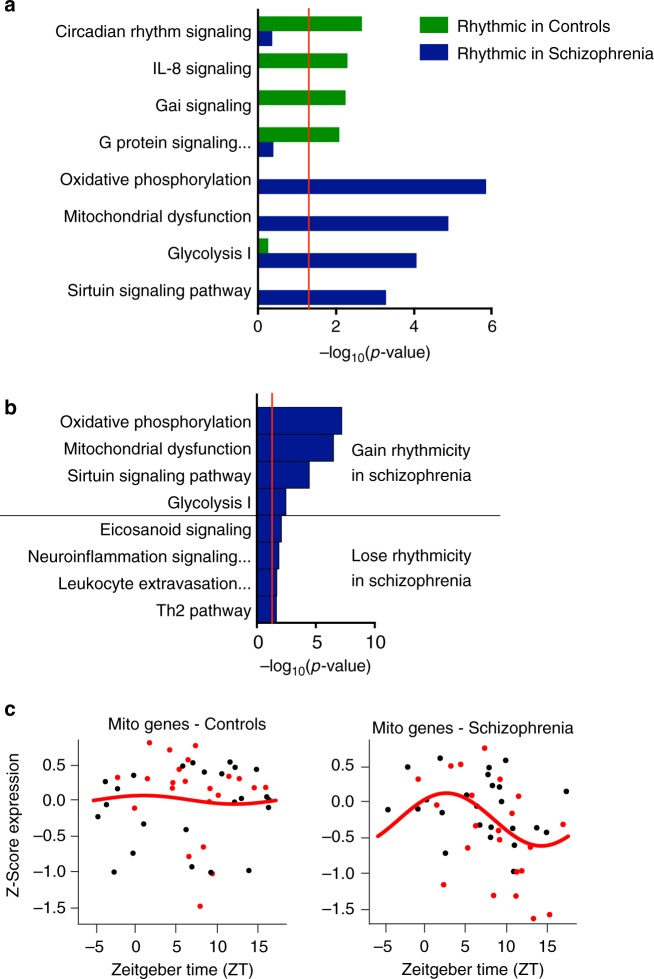
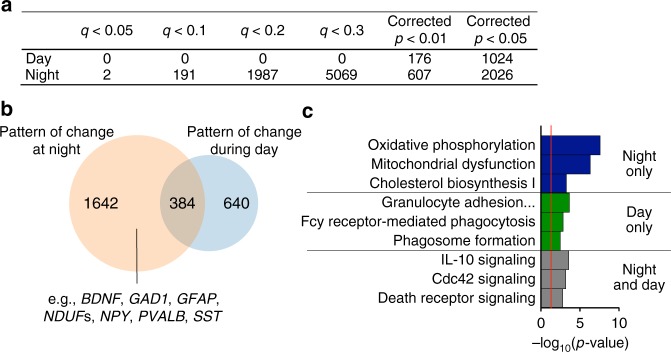
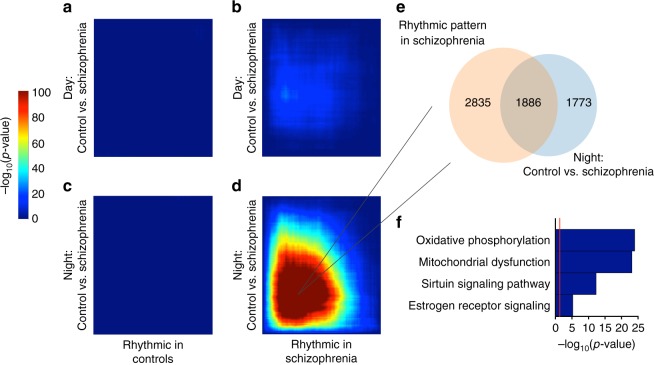
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 MH109897/MH/NIMH NIH HHS/United States
- R37 MH057881/MH/NIMH NIH HHS/United States
- R01 MH075916/MH/NIMH NIH HHS/United States
- U01 MH103392/MH/NIMH NIH HHS/United States
- R01 MH109677/MH/NIMH NIH HHS/United States
- P50 MH084053/MH/NIMH NIH HHS/United States
- HHSN271201300031C/MH/NIMH NIH HHS/United States
- R01 MH110921/MH/NIMH NIH HHS/United States
- P50 MH066392/MH/NIMH NIH HHS/United States
- R01 MH080405/MH/NIMH NIH HHS/United States
- R01 MH085542/MH/NIMH NIH HHS/United States
- R01 MH111601/MH/NIMH NIH HHS/United States
- R01 MH097276/MH/NIMH NIH HHS/United States
- P01 AG002219/AG/NIA NIH HHS/United States
- R01 MH093725/MH/NIMH NIH HHS/United States
- P50 AG005138/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
