

Variable impact of the hemagglutinin polybasic cleavage site on virulence and pathogenesis of avian influenza H7N7 virus in chickens, turkeys and ducks
- PMID: 31399610
- PMCID: PMC6689016
- DOI: 10.1038/s41598-019-47938-3
Variable impact of the hemagglutinin polybasic cleavage site on virulence and pathogenesis of avian influenza H7N7 virus in chickens, turkeys and ducks
Abstract
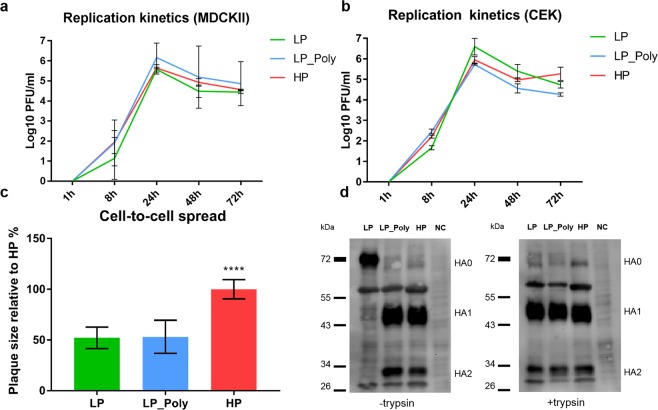
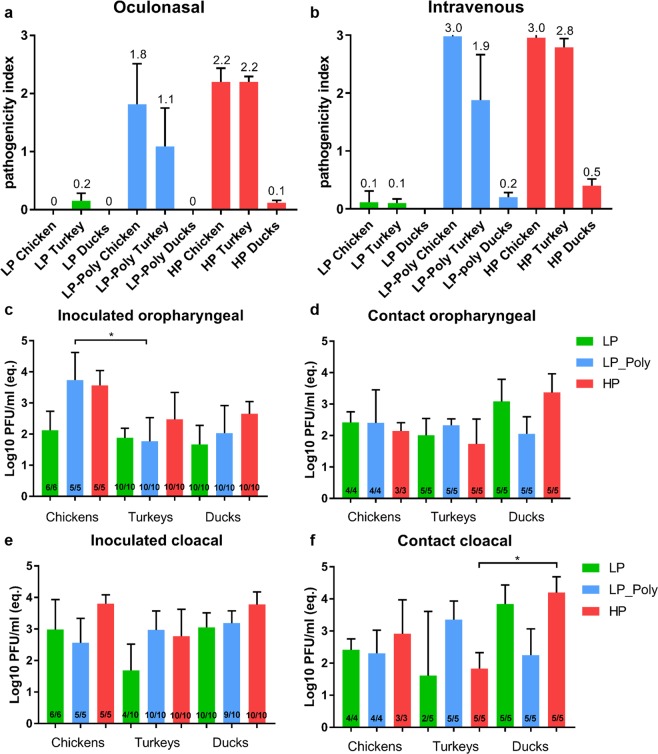
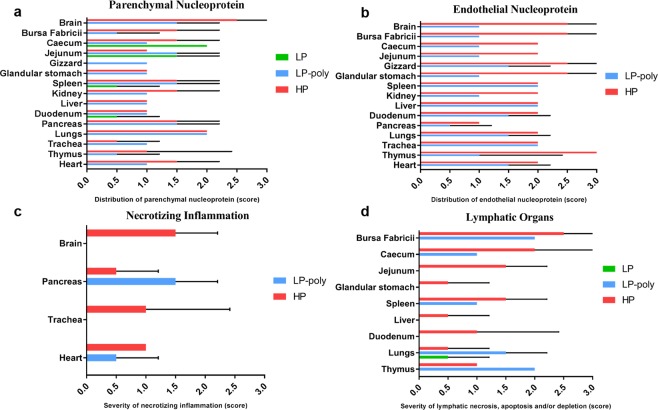
Avian influenza viruses (AIV) are classified into 16 hemagglutinin (HA; H1-H16) and 9 neuraminidase (NA; N1-N9) subtypes. All AIV are low pathogenic (LP) in birds, but subtypes H5 and H7 AIV can evolve into highly pathogenic (HP) forms. In the last two decades evolution of HPAIV H7 from LPAIV has been frequently reported. However, little is known about the pathogenesis and evolution of HP H7 from LP ancestors particularly, in non-chicken hosts. In 2015, both LP and HP H7N7 AIV were isolated from chickens in two neighbouring farms in Germany. Here, the virulence of these isogenic H7N7 LP, HP and LP virus carrying a polybasic HA cleavage site (HACS) from HP (designated LP-Poly) was studied in chickens, turkeys and different duck breeds. The LP precursor was avirulent in all birds. In contrast, all inoculated and contact chickens and turkeys died after infection with HP. HP infected Pekin and Mallard ducks remained clinically healthy, while Muscovy ducks exhibited moderate depression and excreted viruses at significantly higher amounts. The polybasic HACS increased virulence in a species-specific manner with intravenous pathogenicity indices of 3.0, 1.9 and 0.2 in chickens, turkeys and Muscovy ducks, respectively. Infection of endothelial cells was only observed in chickens. In summary, Pekin and Mallard were more resistant to HPAIV H7N7 than chickens, turkeys and Muscovy ducks. The polybasic HACS was the main determinant for virulence and endotheliotropism of HPAIV H7N7 in chickens, whereas other viral and/or host factors play an essential role in virulence and pathogenesis in turkeys and ducks.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Bosch FX, Garten W, Klenk HD, Rott R. Proteolytic Cleavage of Influenza-Virus Hemagglutinins - Primary Structure of the Connecting Peptide between Ha1 and Ha2 Determines Proteolytic Cleavability and Pathogenicity of Avian Influenza-Viruses. Virology. 1981;113:725–735. doi: 10.1016/0042-6822(81)90201-4. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
