

Dynamic architecture of the Escherichia coli structural maintenance of chromosomes (SMC) complex, MukBEF
- PMID: 31400115
- PMCID: PMC6765140
- DOI: 10.1093/nar/gkz696
Dynamic architecture of the Escherichia coli structural maintenance of chromosomes (SMC) complex, MukBEF
Abstract
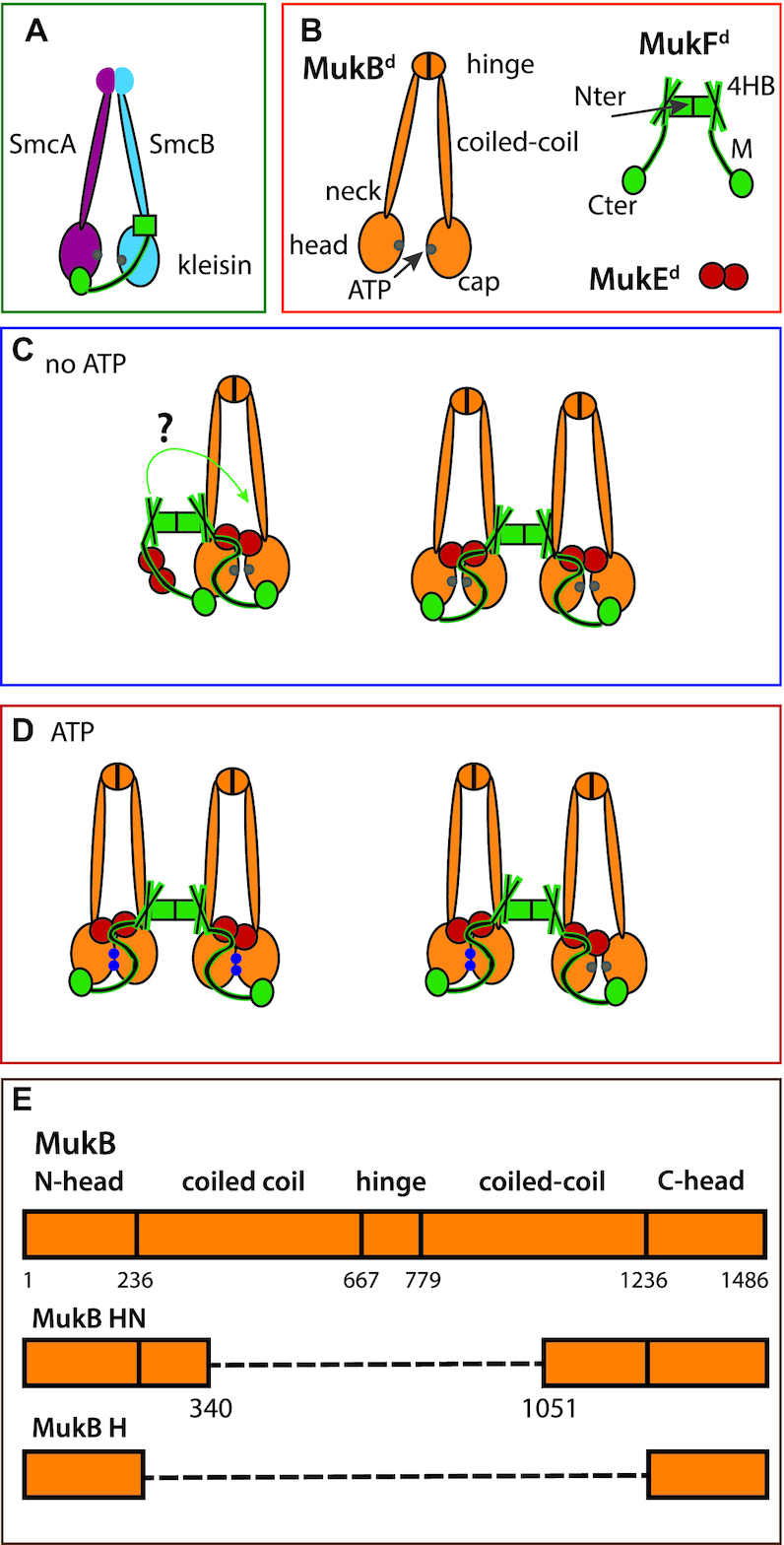
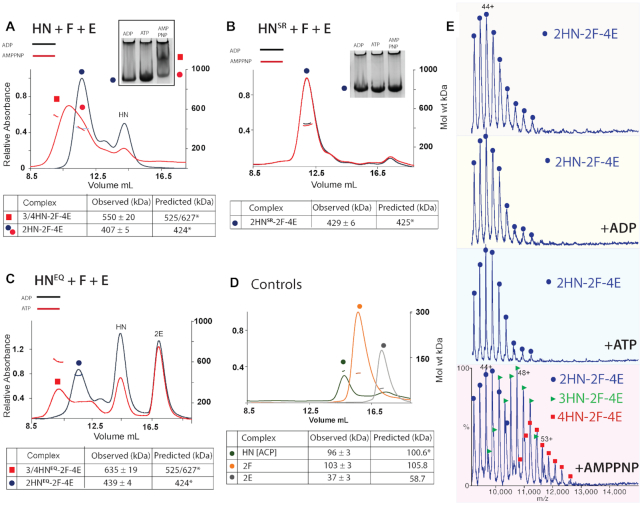
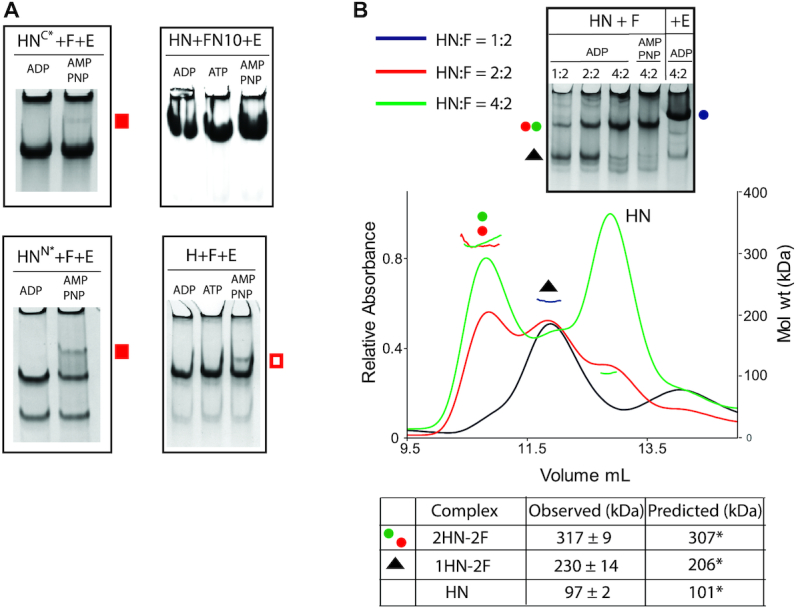
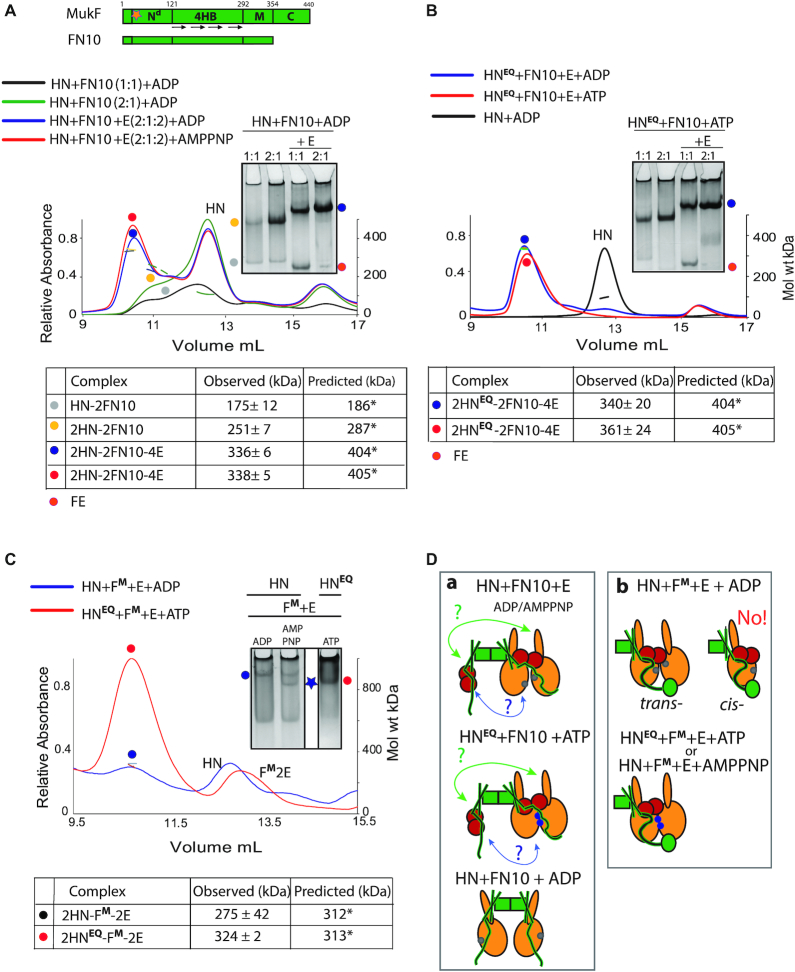
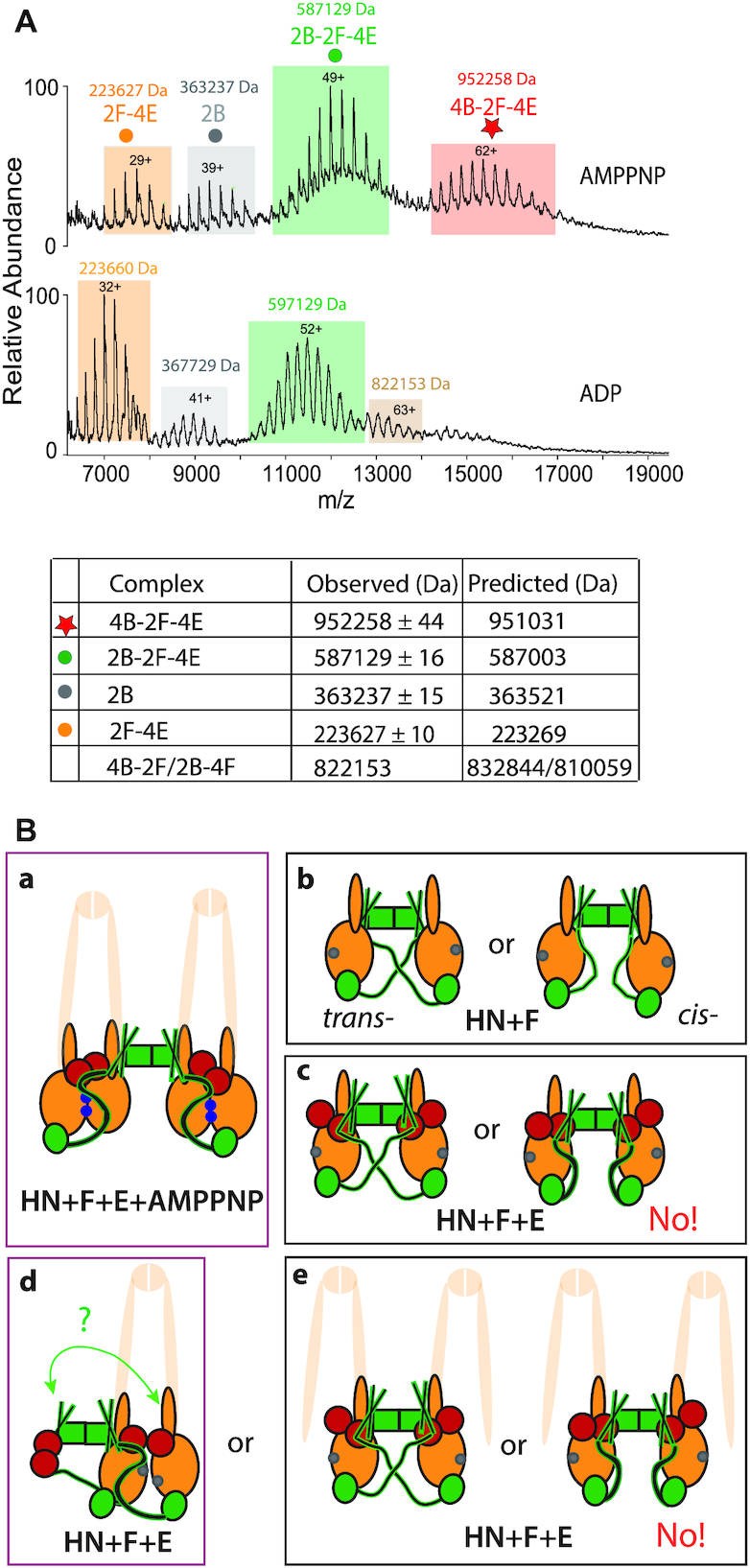
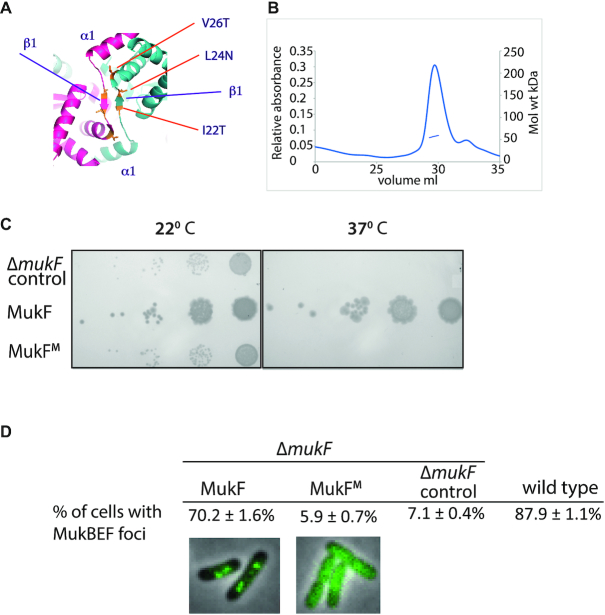
Ubiquitous Structural Maintenance of Chromosomes (SMC) complexes use a proteinaceous ring-shaped architecture to organize and individualize chromosomes, thereby facilitating chromosome segregation. They utilize cycles of adenosine triphosphate (ATP) binding and hydrolysis to transport themselves rapidly with respect to DNA, a process requiring protein conformational changes and multiple DNA contact sites. By analysing changes in the architecture and stoichiometry of the Escherichia coli SMC complex, MukBEF, as a function of nucleotide binding to MukB and subsequent ATP hydrolysis, we demonstrate directly the formation of dimer of MukBEF dimer complexes, dependent on dimeric MukF kleisin. Using truncated and full length MukB, in combination with MukEF, we show that engagement of the MukB ATPase heads on nucleotide binding directs the formation of dimers of heads-engaged dimer complexes. Complex formation requires functional interactions between the C- and N-terminal domains of MukF with the MukB head and neck, respectively, and MukE, which organizes the complexes by stabilizing binding of MukB heads to MukF. In the absence of head engagement, a MukF dimer bound by MukE forms complexes containing only a dimer of MukB. Finally, we demonstrate that cells expressing MukBEF complexes in which MukF is monomeric are Muk-, with the complexes failing to associate with chromosomes.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Bürmann F., Shin H.C., Basquin J., Soh Y.M., Gimenez-Oya V., Kim Y.G., Oh B.H., Gruber S.. An asymmetric SMC-kleisin bridge in prokaryotic condensin. Nat. Struct. Mol. Biol. 2013; 20:371–379. - PubMed
-
- Huis in ’t Veld P.J., Herzog F., Ladurner R., Davidson I.F., Piric S., Kreidl E., Bhaskara V., Aebersold R., Peters J.M.. Characterization of a DNA exit gate in the human cohesin ring. Science. 2014; 346:968–972. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
