

Predictive Sensing: The Role of Motor Signals in Sensory Processing
- PMID: 31401034
- PMCID: PMC6733654
- DOI: 10.1016/j.bpsc.2019.06.003
Predictive Sensing: The Role of Motor Signals in Sensory Processing
Abstract
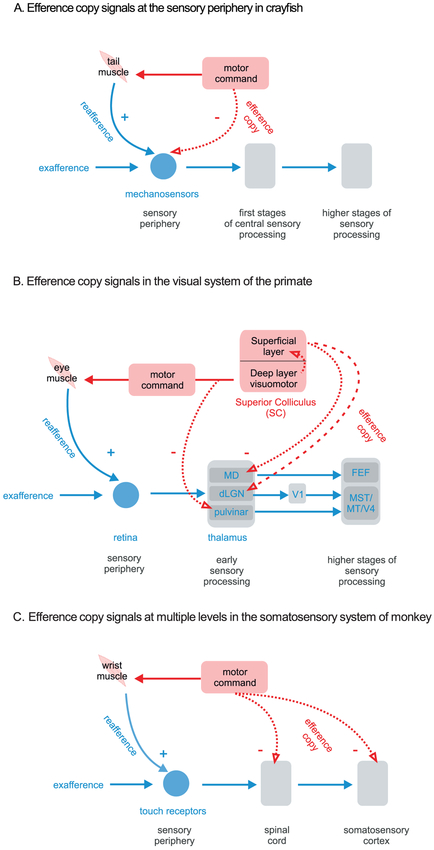
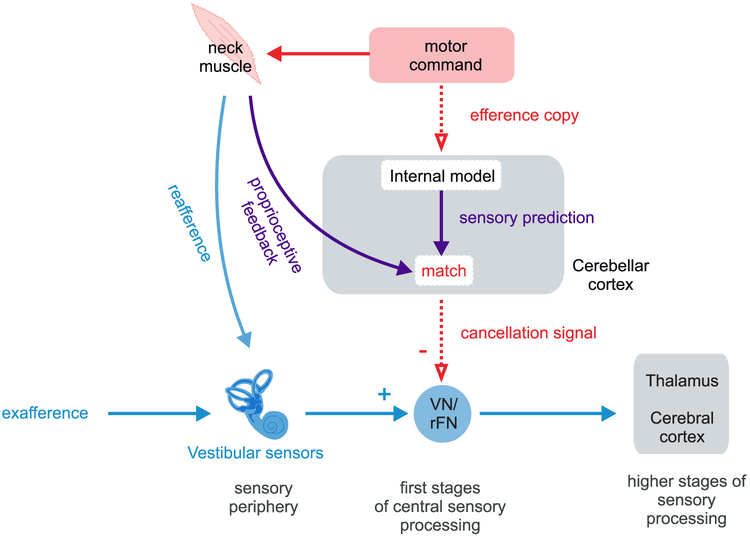
The strategy of integrating motor signals with sensory information during voluntary behavior is a general feature of sensory processing. It is required to distinguish externally applied (exafferent) from self-generated (reafferent) sensory inputs. This distinction, in turn, underlies our ability to achieve both perceptual stability and accurate motor control during everyday activities. In this review, we consider the results of recent experiments that have provided circuit-level insight into how motor-related inputs to sensory areas selectively cancel self-generated sensory inputs during active behaviors. These studies have revealed both common strategies and important differences across systems. Sensory reafference is suppressed at the earliest stages of central processing in the somatosensory, vestibular, and auditory systems, with the cerebellum and cerebellum-like structures playing key roles. Furthermore, motor-related inputs can also suppress reafferent responses at higher levels of processing such as the cortex-a strategy preferentially used in visual processing. These recent findings have important implications for understanding how the brain achieves the flexibility required to continuously calibrate relationships between motor signals and the resultant sensory feedback, a computation necessary for our subjective awareness that we control both our actions and their sensory consequences.
Keywords: Active sensing; Cerebellum; Corollary discharge; Efference copy; Internal model; Prediction.
Copyright © 2019 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
References
-
- Ford JM, Mathalon DH (2005): Corollary discharge dysfunction in schizophrenia: can it explain auditory hallucinations? Int J Psychophysiol. 58:179–189. - PubMed
-
- Helmholtz HV (1925): Handbuch der Physiologischen Optik [Treatise on PhysiologicalOptics]. Southall: Edition ed.: JPC.
-
- Sperry RW (1950): Neural basis of the spontaneous optokinetic response produced by visual inversion. J Comp Physiol Psychol. 43:482–489. - PubMed
-
- von Holst E, Mittelstaedt H (1950): Das reafferenzprinzip. Naturwissenschaften. 37:464–476.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
