

Artery-Associated Sympathetic Innervation Drives Rhythmic Vascular Inflammation of Arteries and Veins
- PMID: 31401849
- PMCID: PMC6756975
- DOI: 10.1161/CIRCULATIONAHA.119.040232
Artery-Associated Sympathetic Innervation Drives Rhythmic Vascular Inflammation of Arteries and Veins
Abstract
Background: The incidence of acute cardiovascular complications is highly time-of-day dependent. However, the mechanisms driving rhythmicity of ischemic vascular events are unknown. Although enhanced numbers of leukocytes have been linked to an increased risk of cardiovascular complications, the role that rhythmic leukocyte adhesion plays in different vascular beds has not been studied.
Methods: We evaluated leukocyte recruitment in vivo by using real-time multichannel fluorescence intravital microscopy of a tumor necrosis factor-α-induced acute inflammation model in both murine arterial and venous macrovasculature and microvasculature. These approaches were complemented with genetic, surgical, and pharmacological ablation of sympathetic nerves or adrenergic receptors to assess their relevance for rhythmic leukocyte adhesion. In addition, we genetically targeted the key circadian clock gene Bmal1 (also known as Arntl) in a lineage-specific manner to dissect the importance of oscillations in leukocytes and components of the vessel wall in this process.
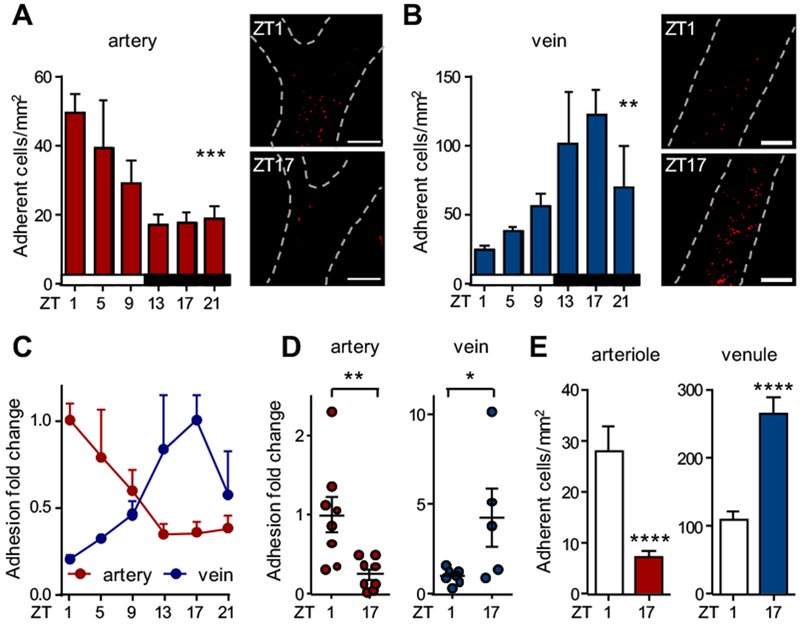
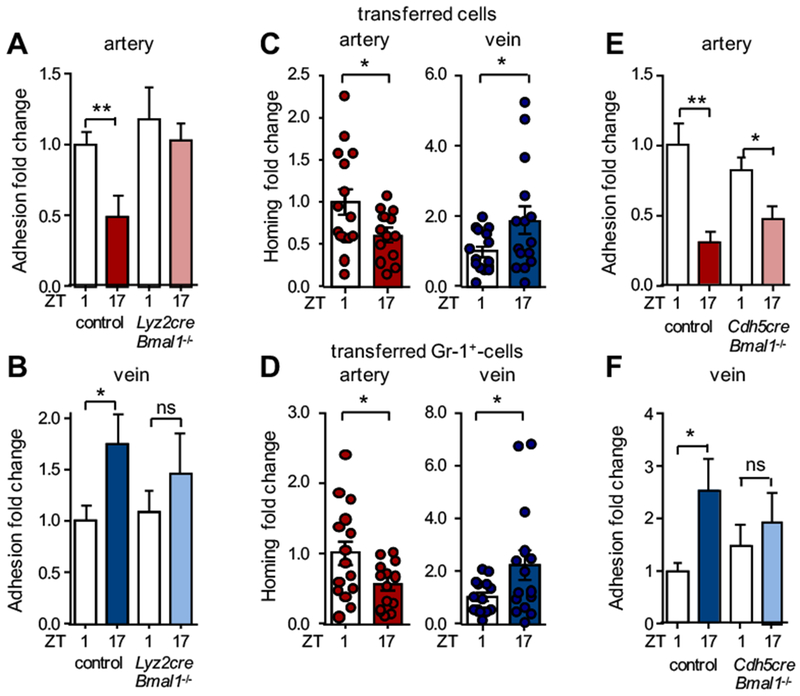
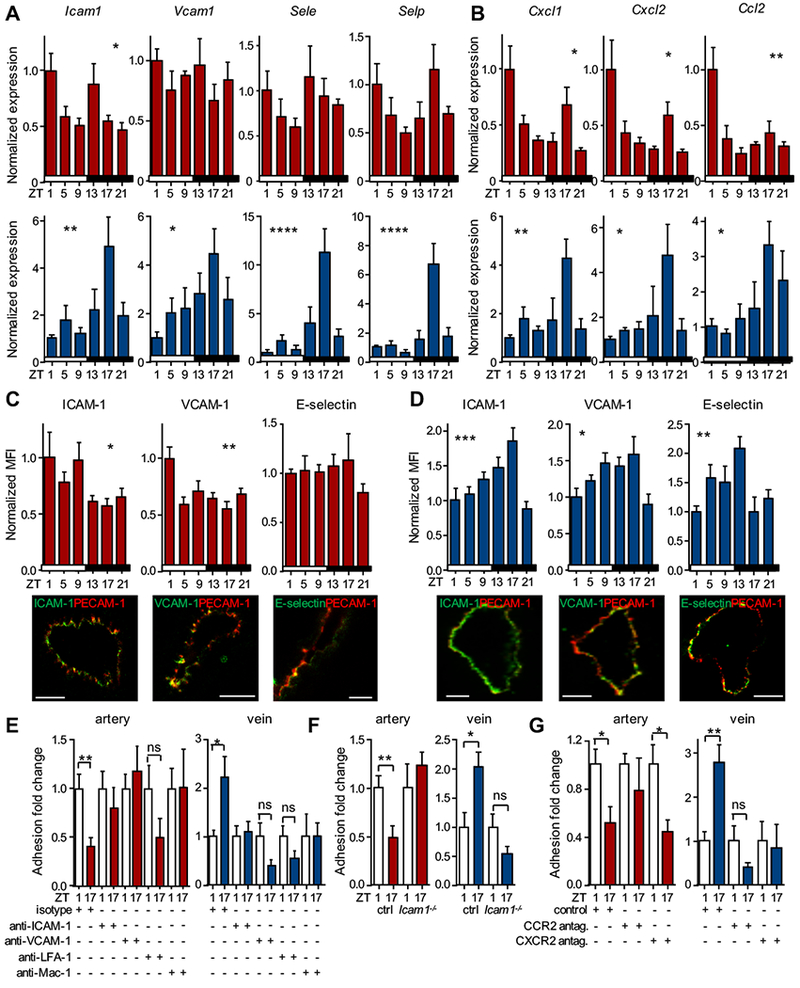
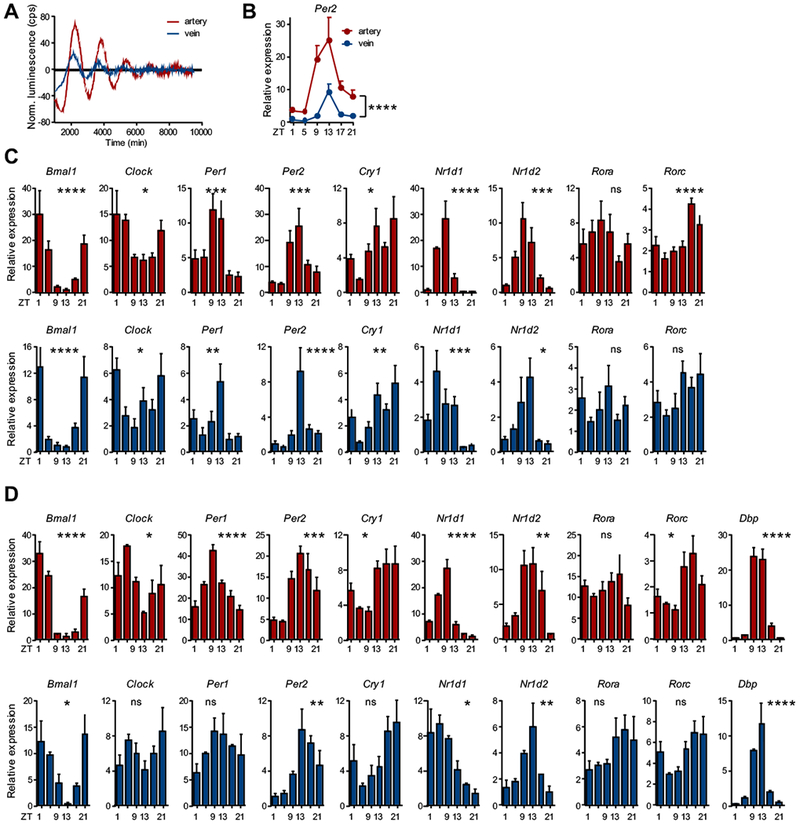
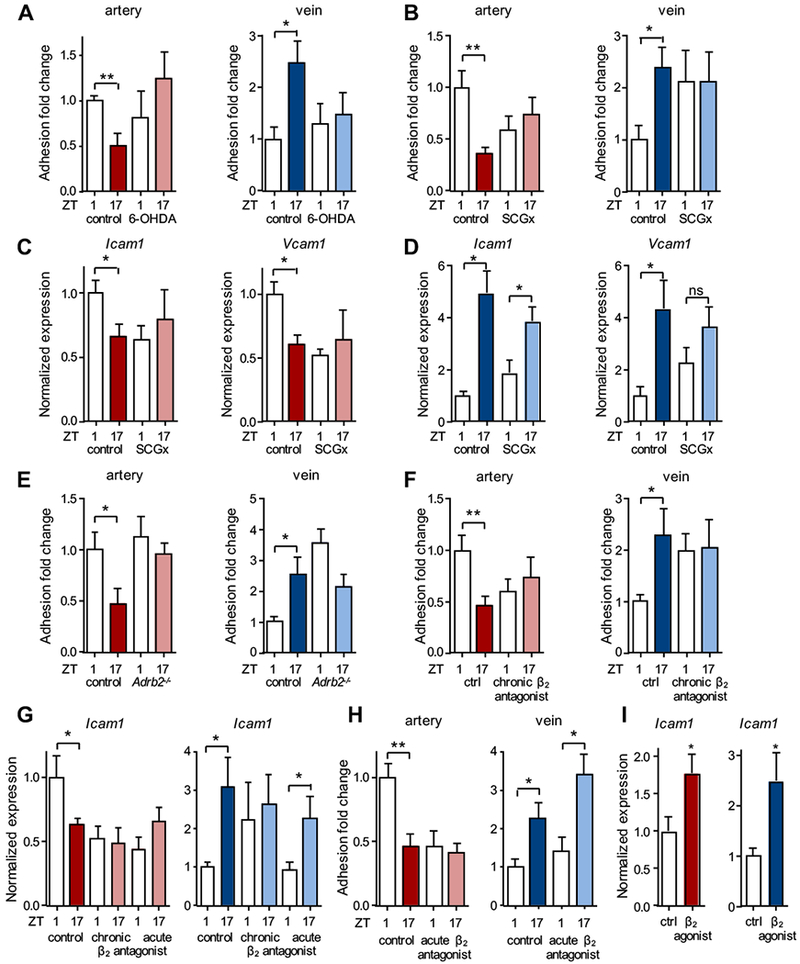
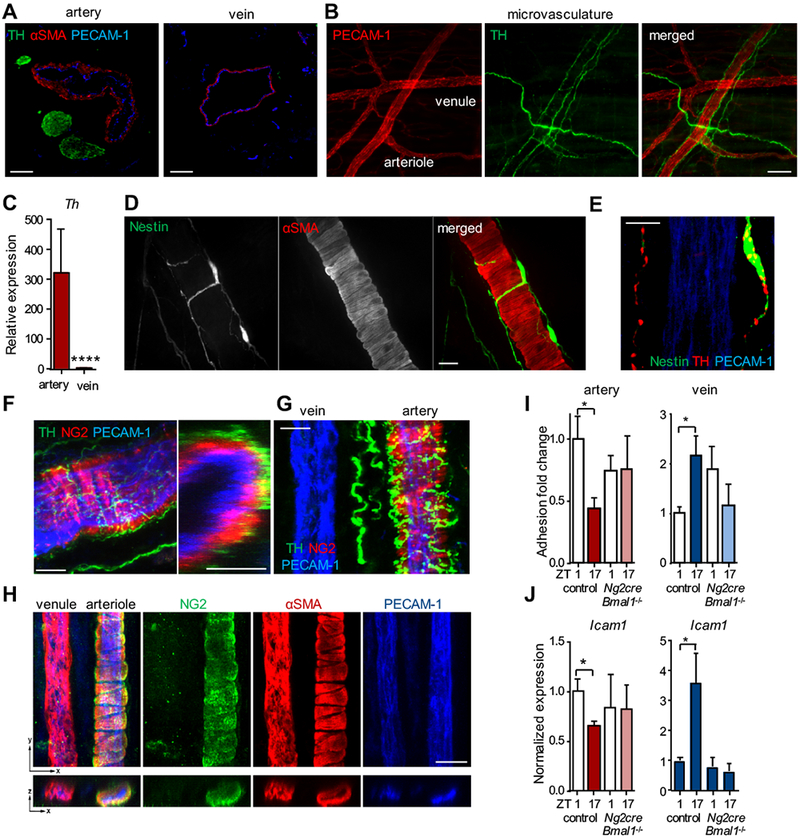
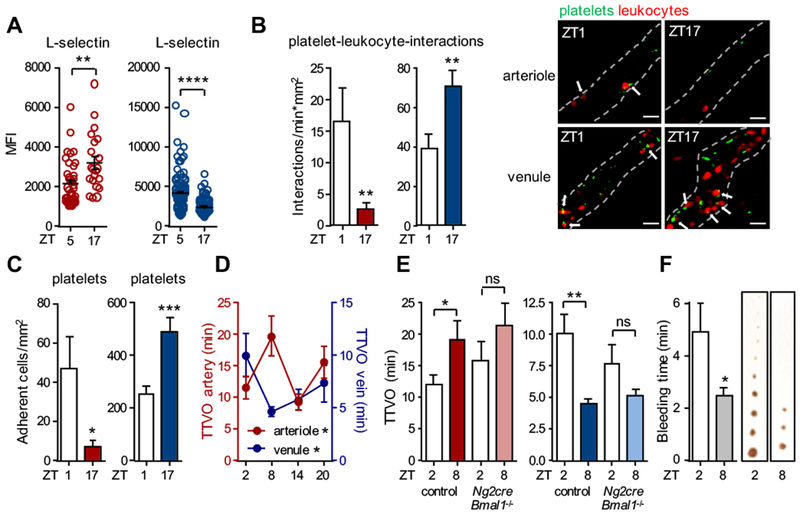
Results: In vivo quantitative imaging analyses of acute inflammation revealed a 24-hour rhythm in leukocyte recruitment to arteries and veins of the mouse macrovasculature and microvasculature. Unexpectedly, although in arteries leukocyte adhesion was highest in the morning, it peaked at night in veins. This phase shift was governed by a rhythmic microenvironment and a vessel type-specific oscillatory pattern in the expression of promigratory molecules. Differences in cell adhesion molecules and leukocyte adhesion were ablated when disrupting sympathetic nerves, demonstrating their critical role in this process and the importance of β2-adrenergic receptor signaling. Loss of the core clock gene Bmal1 in leukocytes, endothelial cells, or arterial mural cells affected the oscillations in a vessel type-specific manner. Rhythmicity in the intravascular reactivity of adherent leukocytes resulted in increased interactions with platelets in the morning in arteries and in veins at night with a higher predisposition to acute thrombosis at different times as a consequence.
Conclusions: Together, our findings point to an important and previously unrecognized role of artery-associated sympathetic innervation in governing rhythmicity in vascular inflammation in both arteries and veins and its potential implications in the occurrence of time-of-day-dependent vessel type-specific thrombotic events.
Keywords: cell adhesion molecules; circadian rhythm; sympathetic nervous system; thrombosis.
Figures
References
-
- Muller JE, Stone PH, Turi ZG, Rutherford JD, Czeisler CA, Parker C, Poole WK, Passamani E, Roberts R, Robertson T, Sobel BE, Willerson JT, and Braunwald E and the MILIS Study Group. Circadian variation in the frequency of onset of acute myocardial infarction. New Eng J MEd. 1985;313:1315–1322. - PubMed
-
- Coller BS. Leukocytosis and ischemic vascular disease morbidity and mortality: is it time to intervene? Arterioscler thromb Vasc Biol. 2005;25:658–670. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
