

Flexible Tethering of ASPP Proteins Facilitates PP-1c Catalysis
- PMID: 31402222
- PMCID: PMC6827562
- DOI: 10.1016/j.str.2019.07.012
Flexible Tethering of ASPP Proteins Facilitates PP-1c Catalysis
Abstract
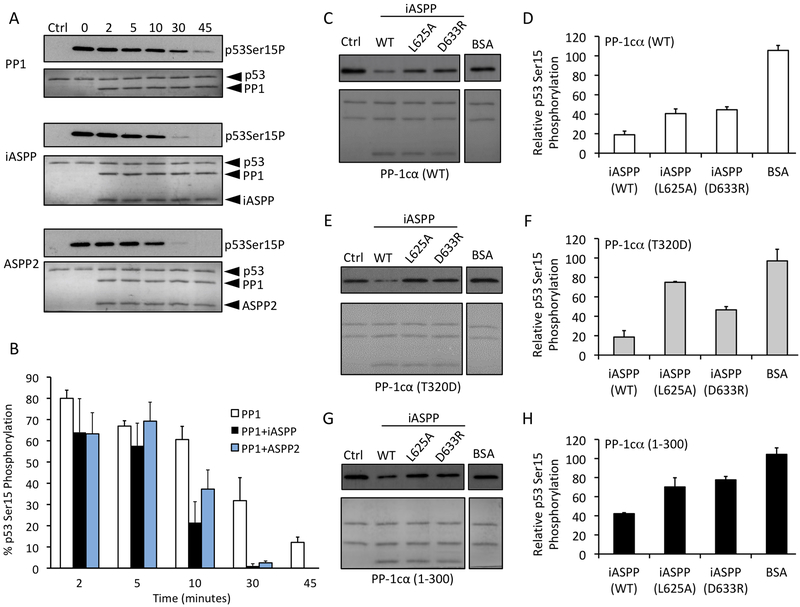
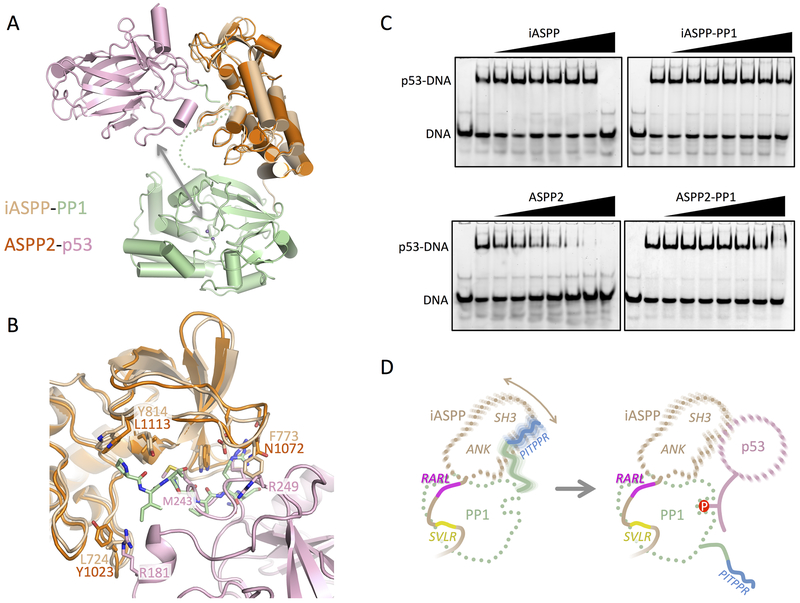
ASPP (apoptosis-stimulating proteins of p53) proteins bind PP-1c (protein phosphatase 1) and regulate p53 impacting cancer cell growth and apoptosis. Here we determine the crystal structure of the oncogenic ASPP protein, iASPP, bound to PP-1c. The structure reveals a 1:1 complex that relies on interactions of the iASPP SILK and RVxF motifs with PP-1c, plus interactions of the PP-1c PxxPxR motif with the iASPP SH3 domain. Small-angle X-ray scattering analyses suggest that the crystal structure undergoes slow interconversion with more extended conformations in solution. We show that iASPP, and the tumor suppressor ASPP2, enhance the catalytic activity of PP-1c against the small-molecule substrate, pNPP as well as p53. The combined results suggest that PxxPxR binding to iASPP SH3 domain is critical for complex formation, and that the modular ASPP-PP-1c interface provides dynamic flexibility that enables functional binding and dephosphorylation of p53 and other diverse protein substrates.
Keywords: ANK repeats; ASPP2; PP-1c; RVxF motif; SH3; Small angle X-ray scattering; X-ray crystallography; dephosphorylation; iASPP; p53.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing financial interests.
Figures
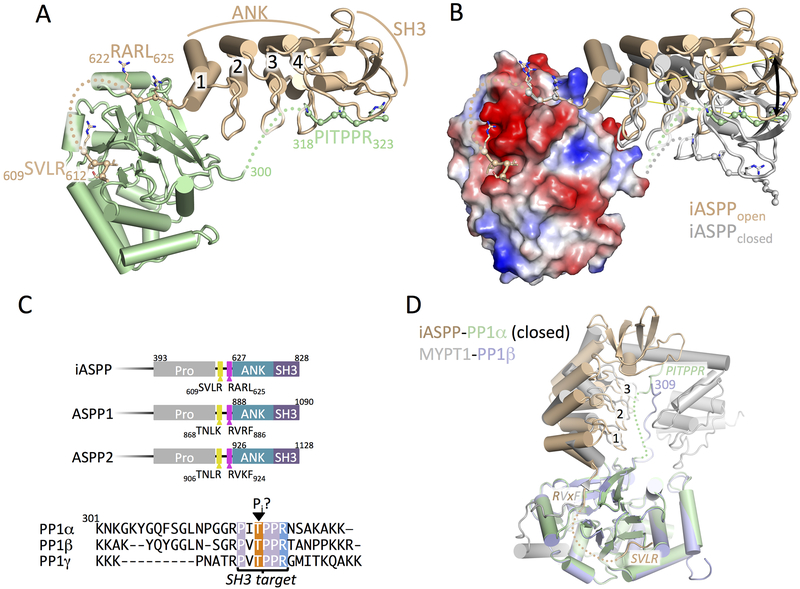
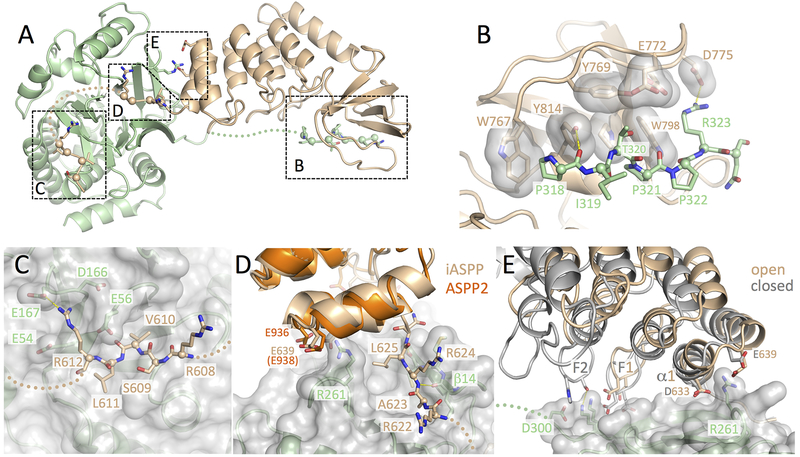
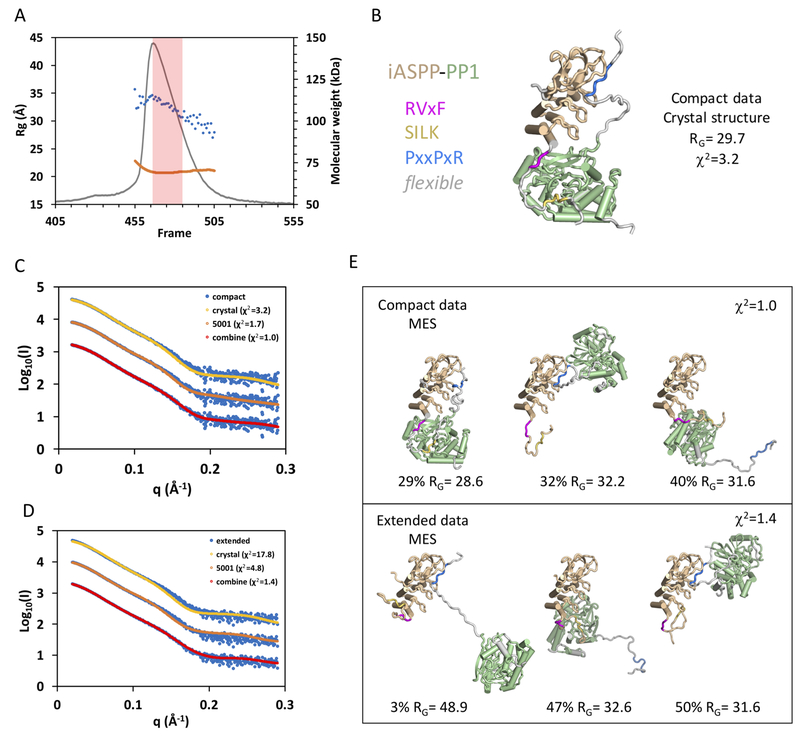
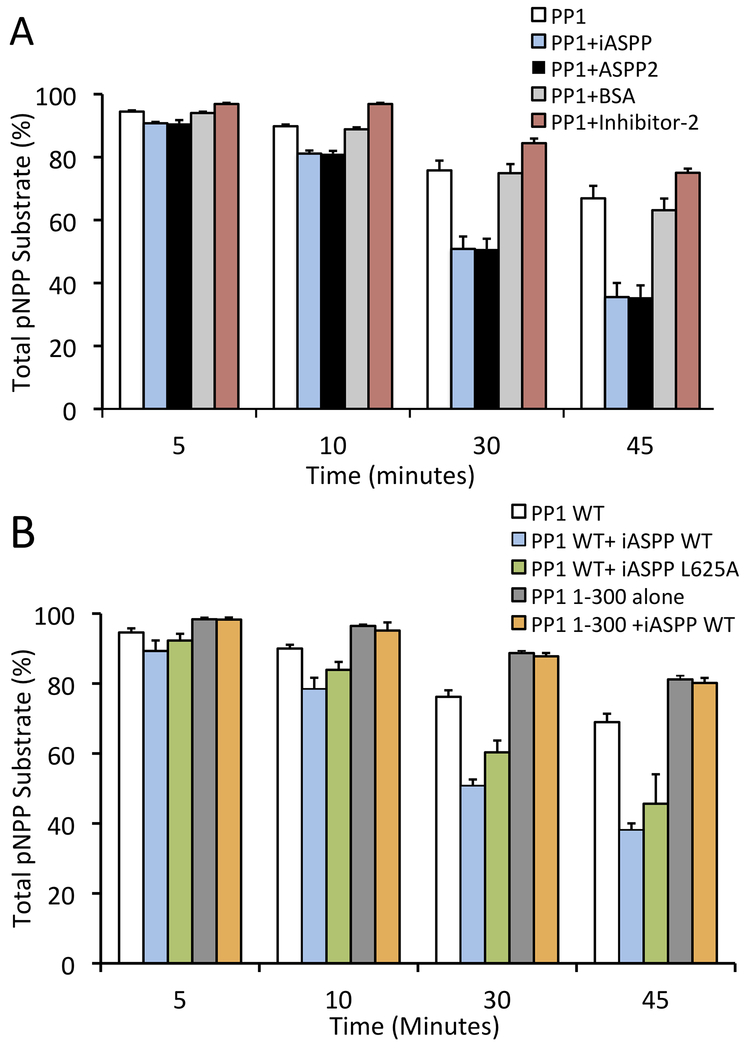
Comment in
-
The More the Merrier: When Four Interfaces Are Better Than One.Structure. 2019 Oct 1;27(10):1483-1484. doi: 10.1016/j.str.2019.09.002. Structure. 2019. PMID: 31577936
References
-
- Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, et al. (2010). PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta crystallographica Section D, Biological crystallography 66, 213–221. - PMC - PubMed
-
- Agirre X, Roman-Gomez J, Jimenez-Velasco A, Garate L, Montiel-Duarte C, Navarro G, Vazquez I, Zalacain M, Calasanz MJ, Heiniger A, et al. (2006). ASPP1, a common activator of TP53, is inactivated by aberrant methylation of its promoter in acute lymphoblastic leukemia. Oncogene 25, 1862–1870. - PubMed
-
- Ao Y, Rohde LH, and Naumovski L (2001). p53-interacting protein 53BP2 inhibits clonogenic survival and sensitizes cells to doxorubicin but not paclitaxel-induced apoptosis. Oncogene 20, 2720–2725. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
