

Energy metabolism controls phenotypes by protein efficiency and allocation
- PMID: 31405984
- PMCID: PMC6717264
- DOI: 10.1073/pnas.1906569116
Energy metabolism controls phenotypes by protein efficiency and allocation
Abstract
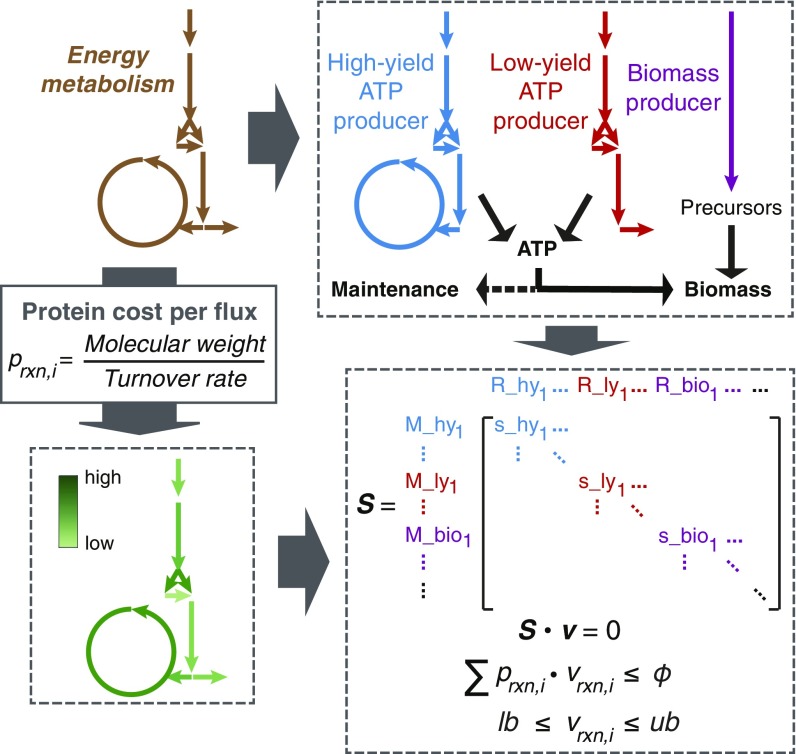
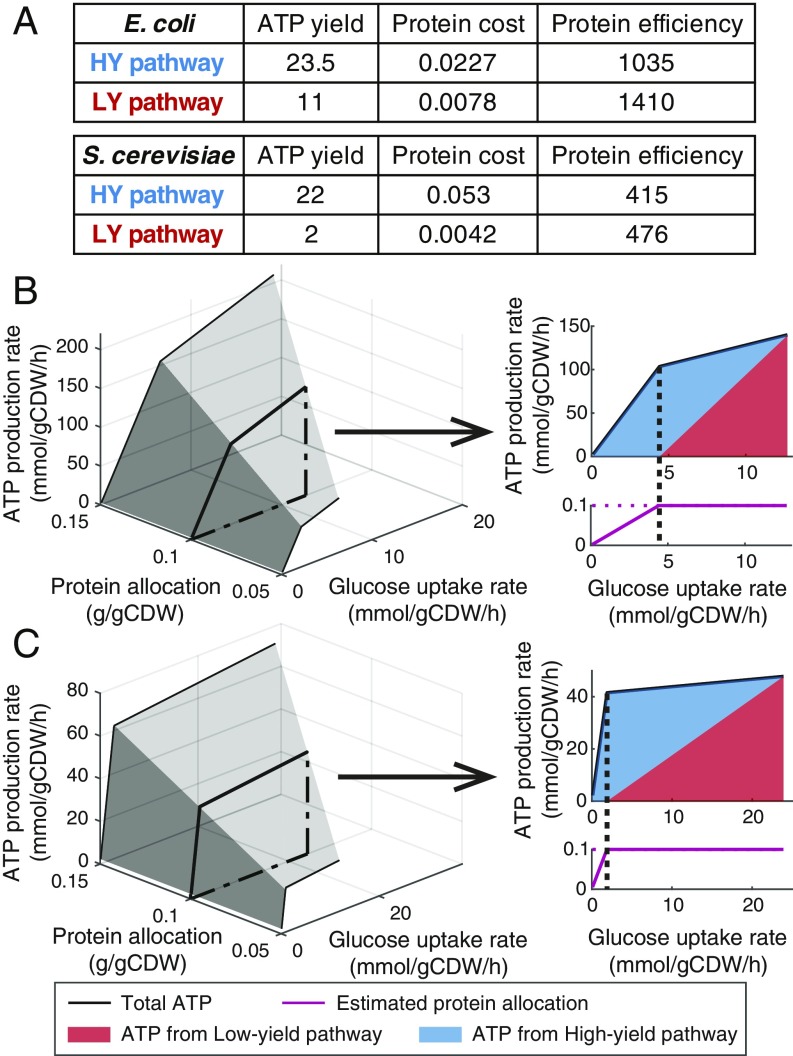
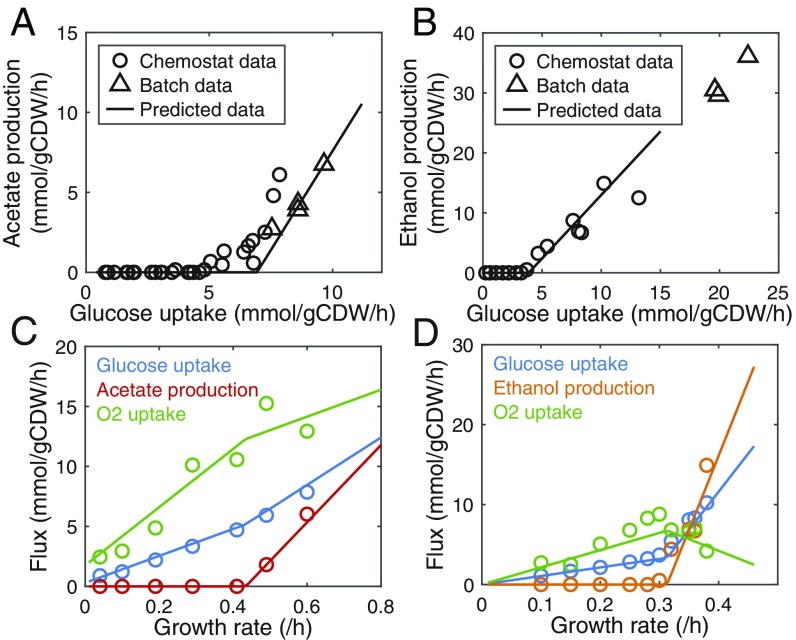
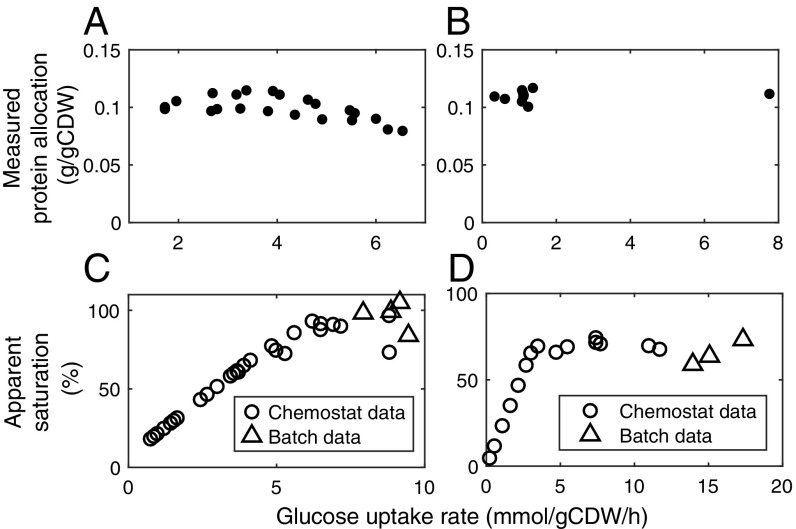
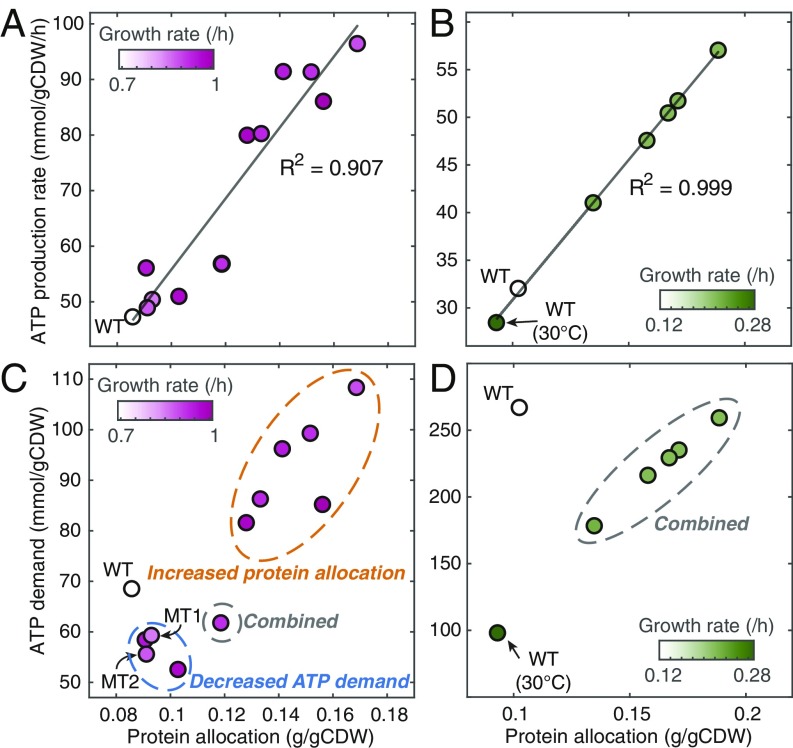
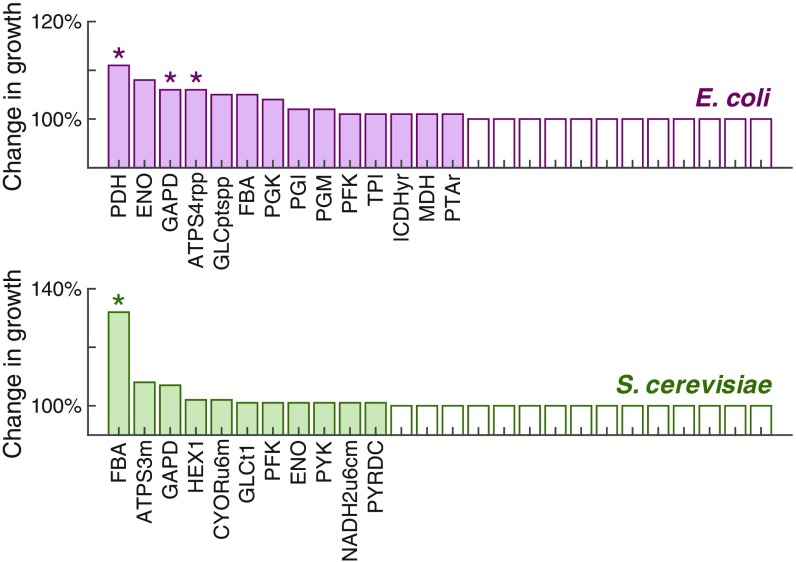
Cells require energy for growth and maintenance and have evolved to have multiple pathways to produce energy in response to varying conditions. A basic question in this context is how cells organize energy metabolism, which is, however, challenging to elucidate due to its complexity, i.e., the energy-producing pathways overlap with each other and even intertwine with biomass formation pathways. Here, we propose a modeling concept that decomposes energy metabolism into biomass formation and ATP-producing pathways. The latter can be further decomposed into a high-yield and a low-yield pathway. This enables independent estimation of protein efficiency for each pathway. With this concept, we modeled energy metabolism for Escherichia coli and Saccharomyces cerevisiae and found that the high-yield pathway shows lower protein efficiency than the low-yield pathway. Taken together with a fixed protein constraint, we predict overflow metabolism in E. coli and the Crabtree effect in S. cerevisiae, meaning that energy metabolism is sufficient to explain the metabolic switches. The static protein constraint is supported by the findings that protein mass of energy metabolism is conserved across conditions based on absolute proteomics data. This also suggests that enzymes may have decreased saturation or activity at low glucose uptake rates. Finally, our analyses point out three ways to improve growth, i.e., increasing protein allocation to energy metabolism, decreasing ATP demand, or increasing activity for key enzymes.
Keywords: Escherichia coli; Saccharomyces cerevisiae; constraint-based modeling; growth rate; metabolic switch.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
