

Cryptic activation of an Irf8 enhancer governs cDC1 fate specification
- PMID: 31406378
- PMCID: PMC6707878
- DOI: 10.1038/s41590-019-0450-x
Cryptic activation of an Irf8 enhancer governs cDC1 fate specification
Abstract
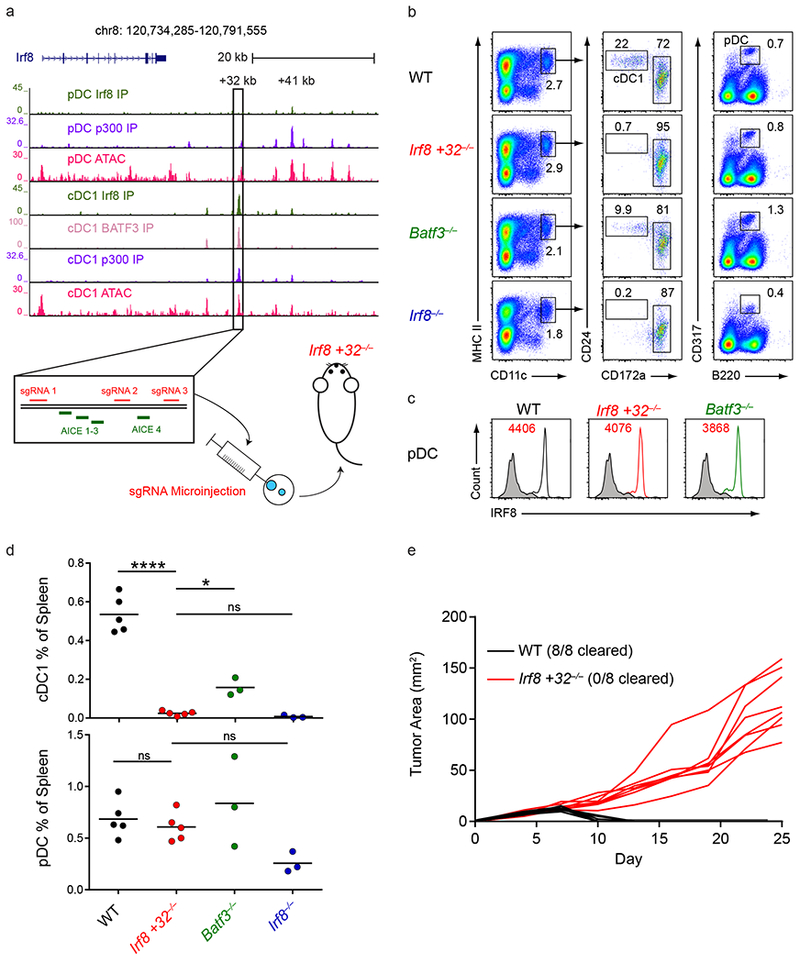
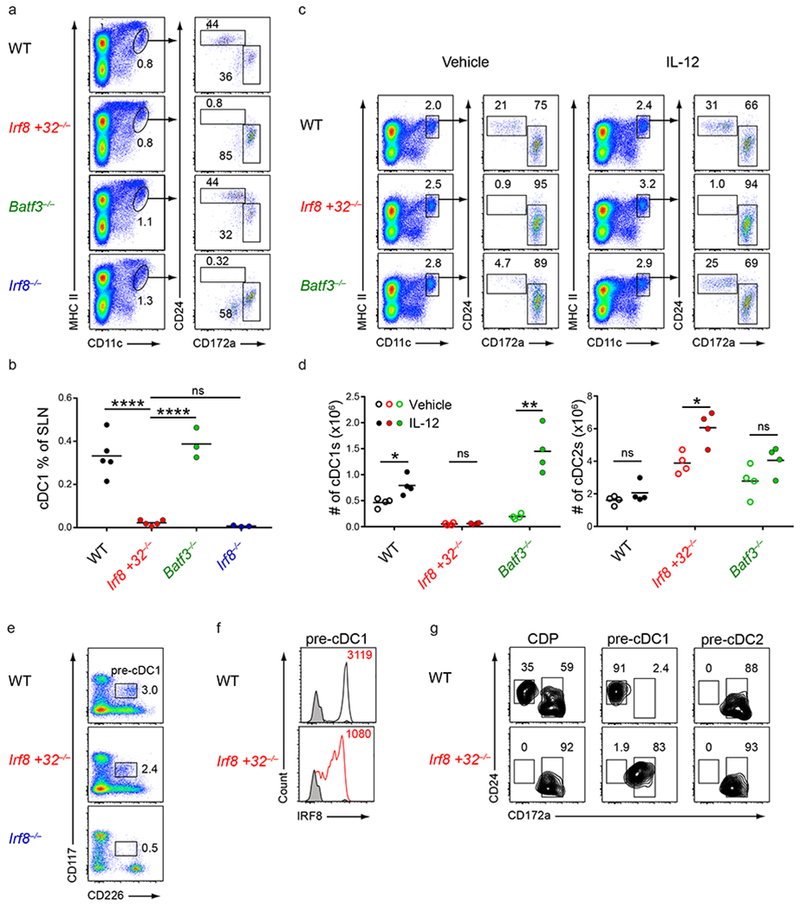
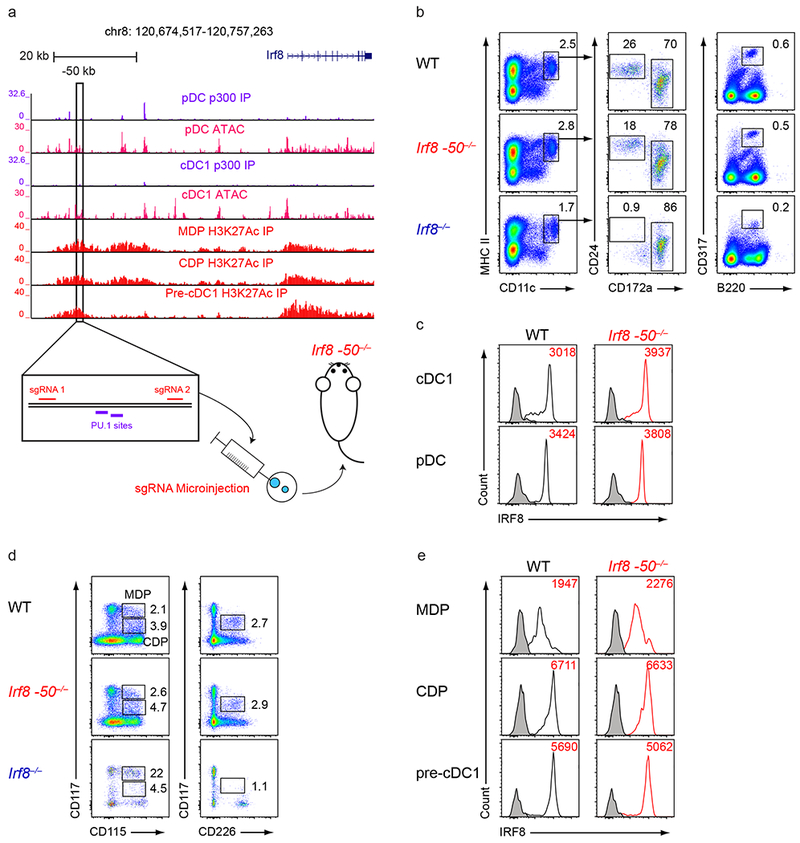
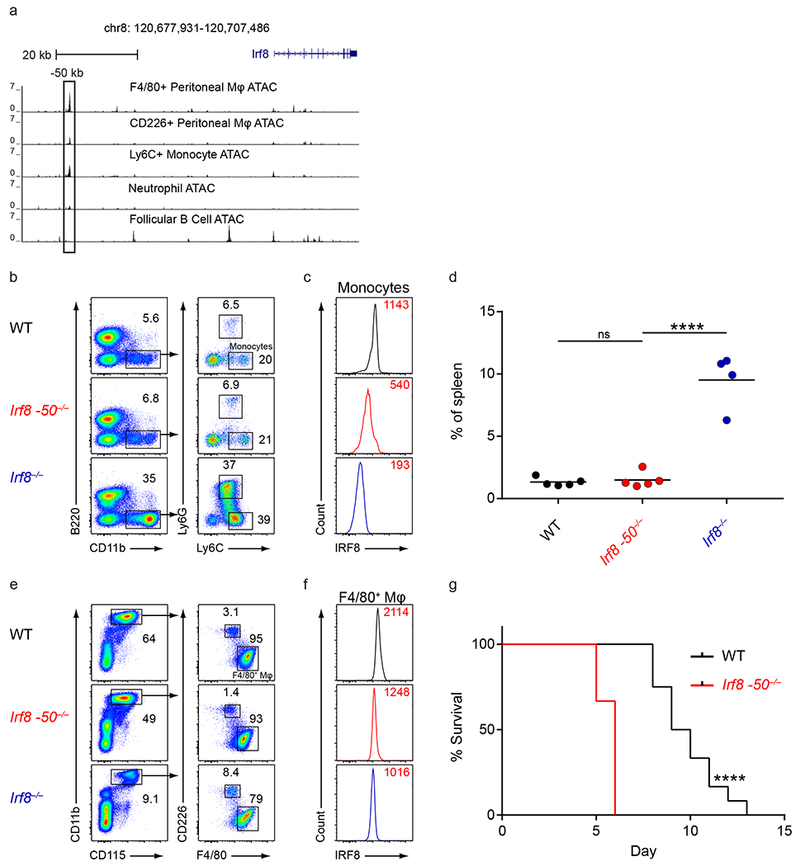
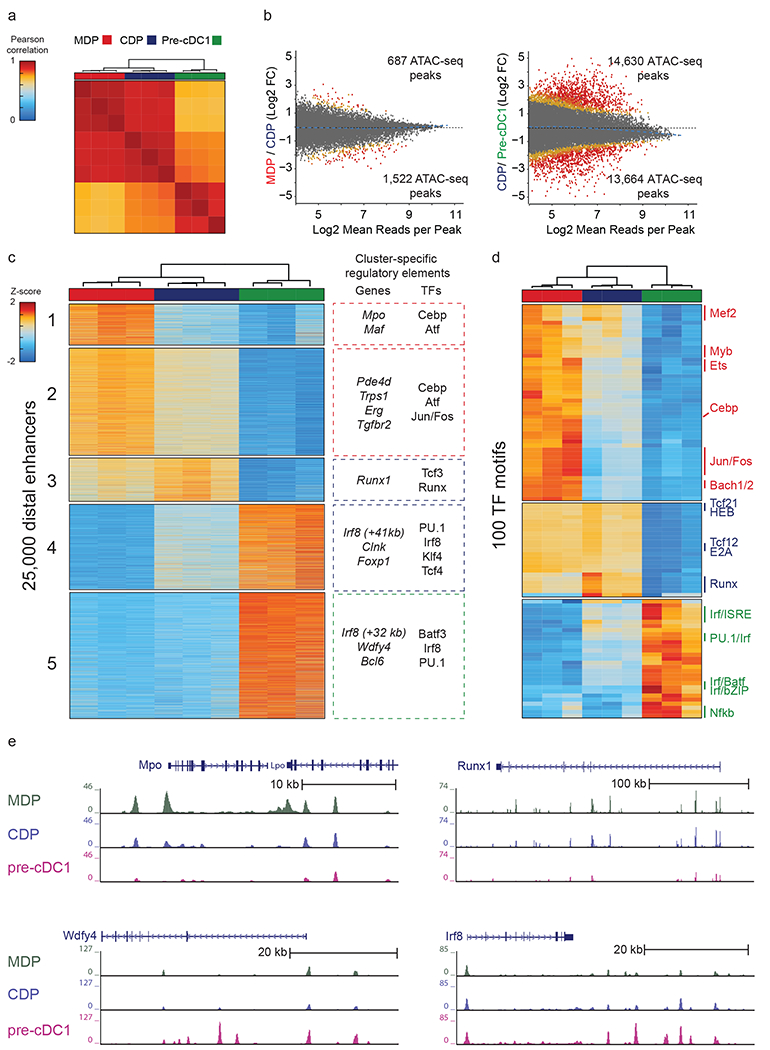
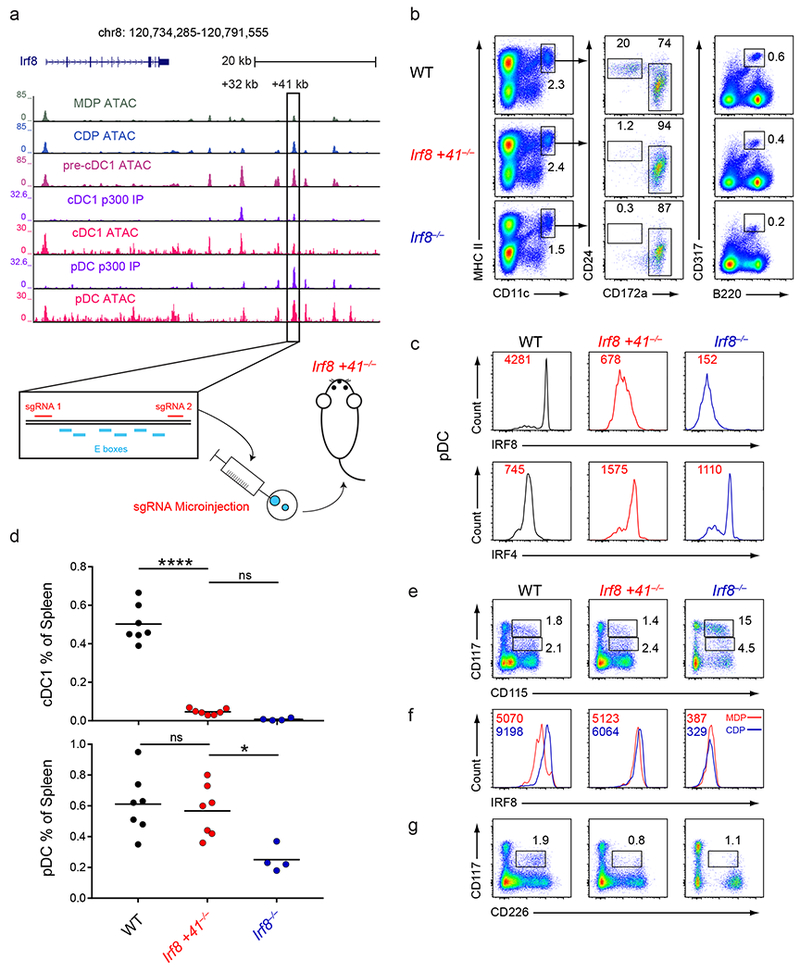
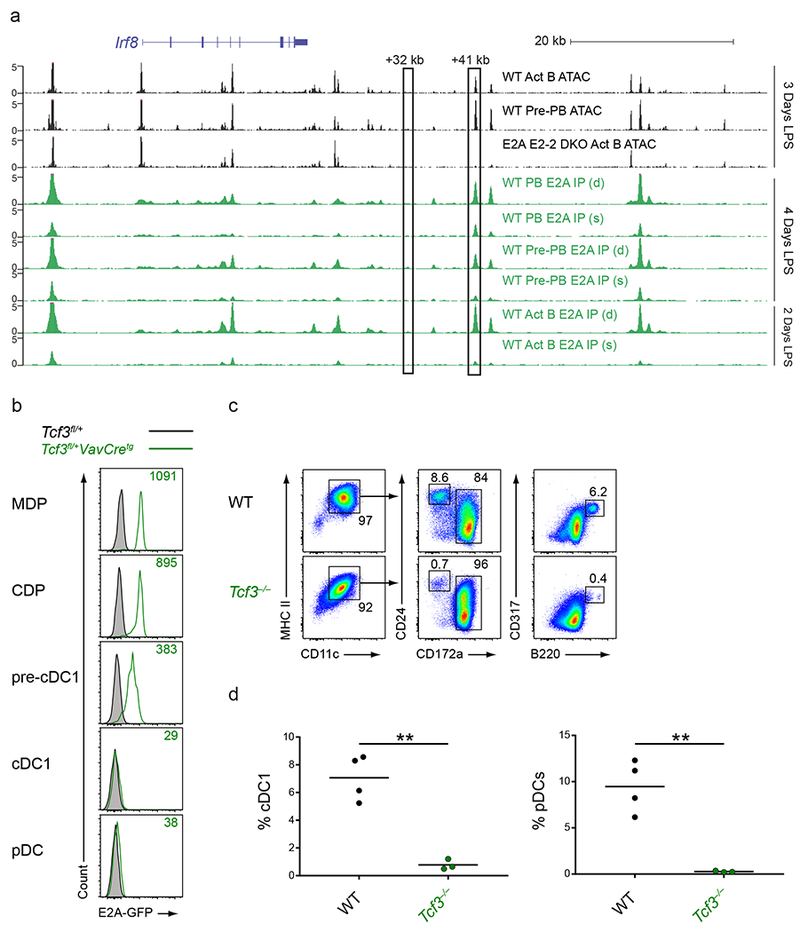
Induction of the transcription factor Irf8 in the common dendritic cell progenitor (CDP) is required for classical type 1 dendritic cell (cDC1) fate specification, but the mechanisms controlling this induction are unclear. In the present study Irf8 enhancers were identified via chromatin profiling of dendritic cells and CRISPR/Cas9 genome editing was used to assess their roles in Irf8 regulation. An enhancer 32 kilobases (kb) downstream of the Irf8 transcriptional start site (+32-kb Irf8) that was active in mature cDC1s was required for the development of this lineage, but not for its specification. Instead, a +41-kb Irf8 enhancer, previously thought to be active only in plasmacytoid dendritic cells, was found to also be transiently accessible in cDC1 progenitors, and deleting this enhancer prevented the induction of Irf8 in CDPs and abolished cDC1 specification. Thus, cryptic activation of the +41-kb Irf8 enhancer in dendritic cell progenitors is responsible for cDC1 fate specification.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
References
-
- Graf T and Enver T Forcing cells to change lineages. Nature 462, 587–594 (2009). - PubMed
-
- Miyamoto T et al. , Myeloid or lymphoid promiscuity as a critical step in hematopoietic lineage commitment. Dev.Cell 3, 137–147 (2002). - PubMed
-
- Hoppe PS et al. , Early myeloid lineage choice is not initiated by random PU.1 to GATA1 protein ratios. Nature 535, 299–302 (2016). - PubMed
Methods-only References
-
- Ranganath S et al. , GATA-3-dependent enhancer activity in IL-4 gene regulation. Journal of Immunology 161, 3822–3826 (1998). - PubMed
-
- Sedy JR et al. , B and T lymphocyte attenuator regulates T cell activation through interaction with herpesvirus entry mediator. Nat.Immunol . 6, 90–98 (2005). - PubMed
-
- Zhu H et al. , Unexpected characteristics of the IFN-gamma reporters in nontransformed T cells. Journal of Immunology 167, 855–865 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI150297/AI/NIAID NIH HHS/United States
- R01 DK097317/DK/NIDDK NIH HHS/United States
- P50 HG007735/HG/NHGRI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- K08 CA230188/CA/NCI NIH HHS/United States
- P30 CA091842/CA/NCI NIH HHS/United States
- UL1 TR000448/TR/NCATS NIH HHS/United States
- R01 AI106352/AI/NIAID NIH HHS/United States
- UL1 TR002345/TR/NCATS NIH HHS/United States
- R01 CA248919/CA/NCI NIH HHS/United States
- R01 AI076427/AI/NIAID NIH HHS/United States
- P30 AR073752/AR/NIAMS NIH HHS/United States
- R01 AI162643/AI/NIAID NIH HHS/United States
- F30 DK108498/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
