

Connecting empirical phenomena and theoretical models of biological coordination across scales
- PMID: 31409241
- PMCID: PMC6731488
- DOI: 10.1098/rsif.2019.0360
Connecting empirical phenomena and theoretical models of biological coordination across scales
Abstract
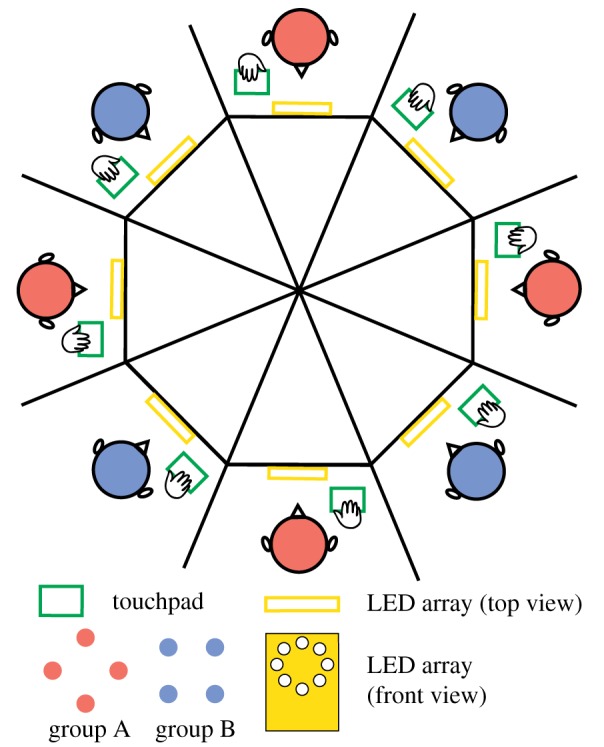
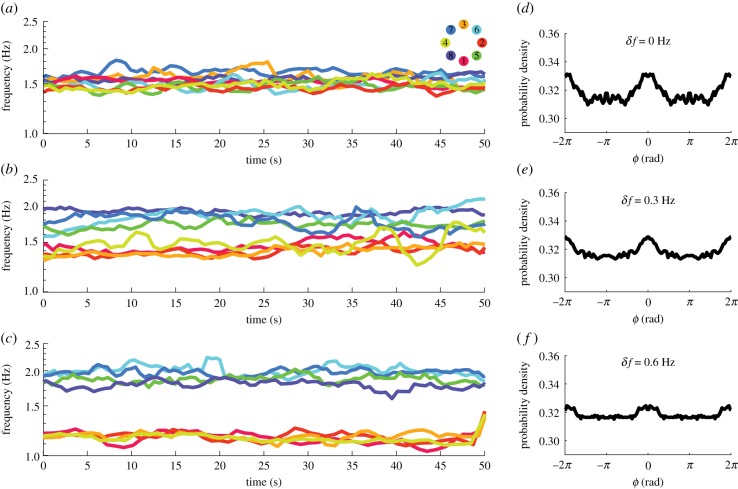
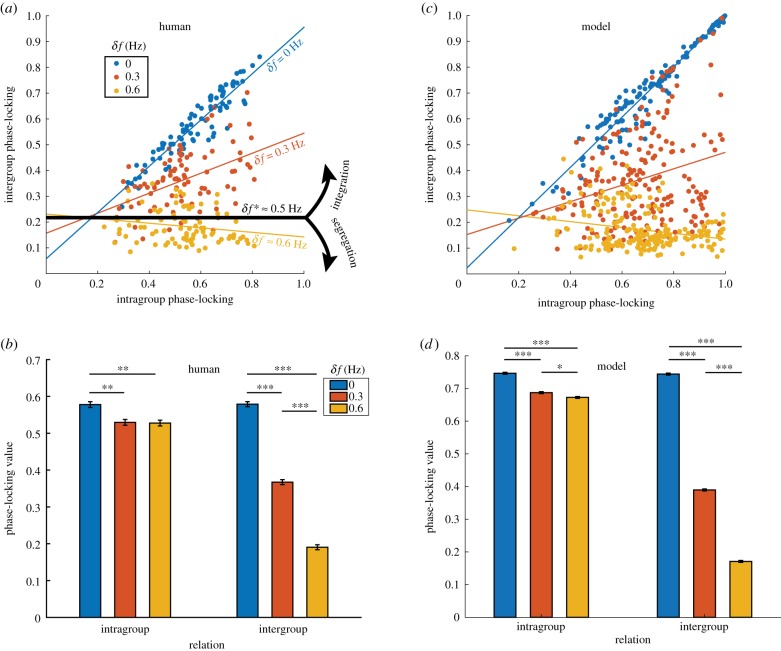
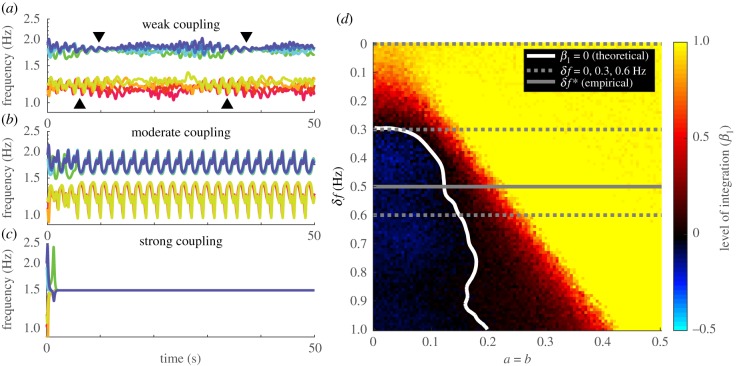
Coordination in living systems-from cells to people-must be understood at multiple levels of description. Analyses and modelling of empirically observed patterns of biological coordination often focus either on ensemble-level statistics in large-scale systems with many components, or on detailed dynamics in small-scale systems with few components. The two approaches have proceeded largely independent of each other. To bridge this gap between levels and scales, we have recently conducted a human experiment of mid-scale social coordination specifically designed to reveal coordination at multiple levels (ensemble, subgroups and dyads) simultaneously. Based on this experiment, the present work shows that, surprisingly, a single system of equations captures key observations at all relevant levels. It also connects empirically validated models of large- and small-scale biological coordination-the Kuramoto and extended Haken-Kelso-Bunz (HKB) models-and the hallmark phenomena that each is known to capture. For example, it exhibits both multistability and metastability observed in small-scale empirical research (via the second-order coupling and symmetry breaking in extended HKB) and the growth of biological complexity as a function of scale (via the scalability of the Kuramoto model). Only by incorporating both of these features simultaneously can we reproduce the essential coordination behaviour observed in our experiment.
Keywords: complex systems; complexity; coordination dynamics; nonlinear dynamics; social; statistical mechanics.
Figures
References
-
- Winfree AT. 1987. The timing of biological clocks. New York, NY: Scientific American Books.
-
- Kuramoto Y. 1984. Chemical oscillations, waves, and turbulence, vol. 19 of Springer Series in Synergetics. Berlin, Germany: Springer.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous