

Gulp1 controls Eph/ephrin trogocytosis and is important for cell rearrangements during development
- PMID: 31409653
- PMCID: PMC6781437
- DOI: 10.1083/jcb.201901032
Gulp1 controls Eph/ephrin trogocytosis and is important for cell rearrangements during development
Abstract
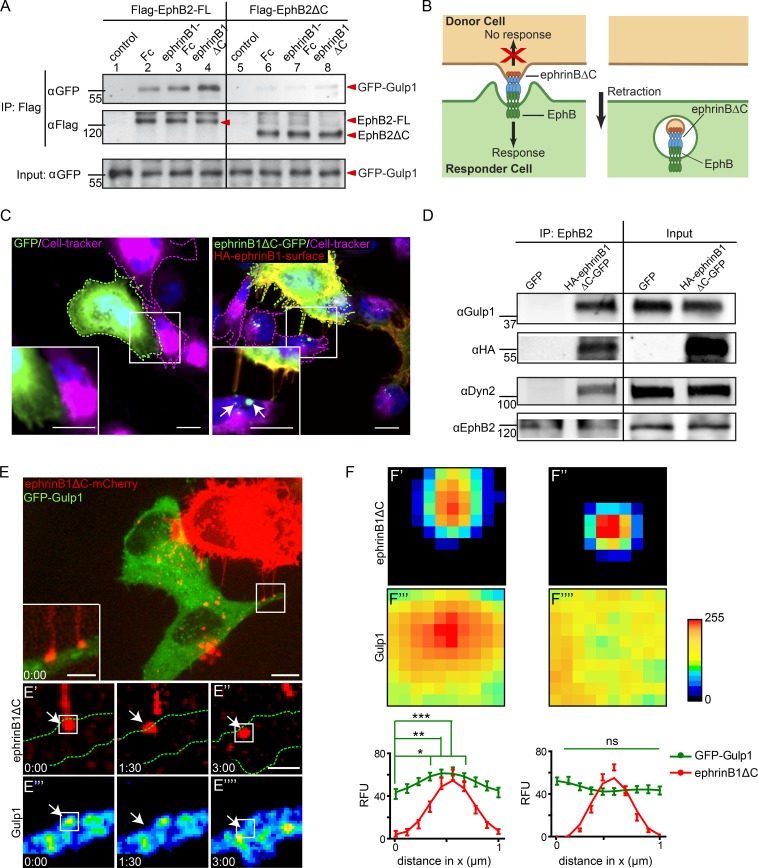
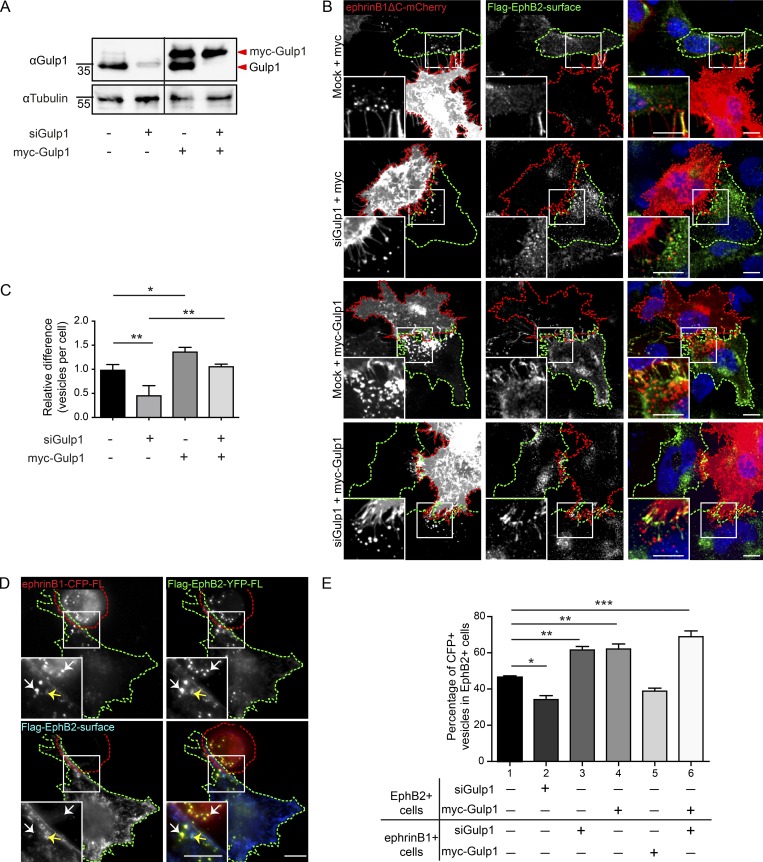
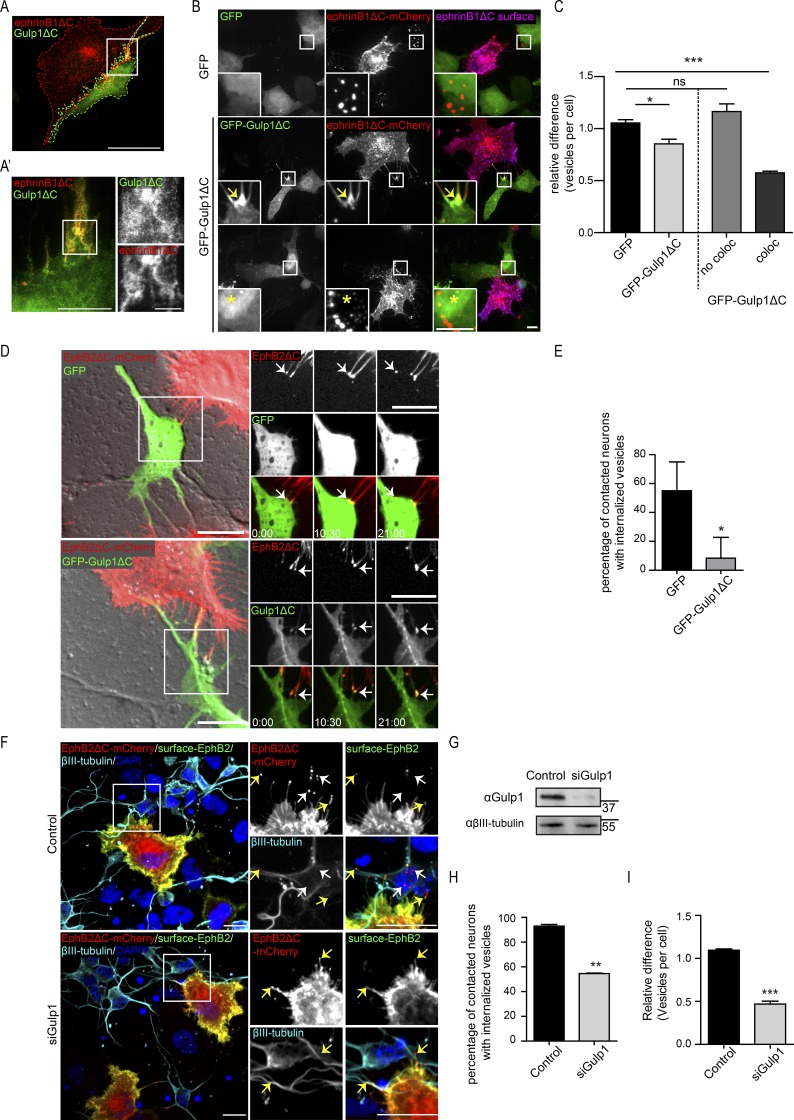
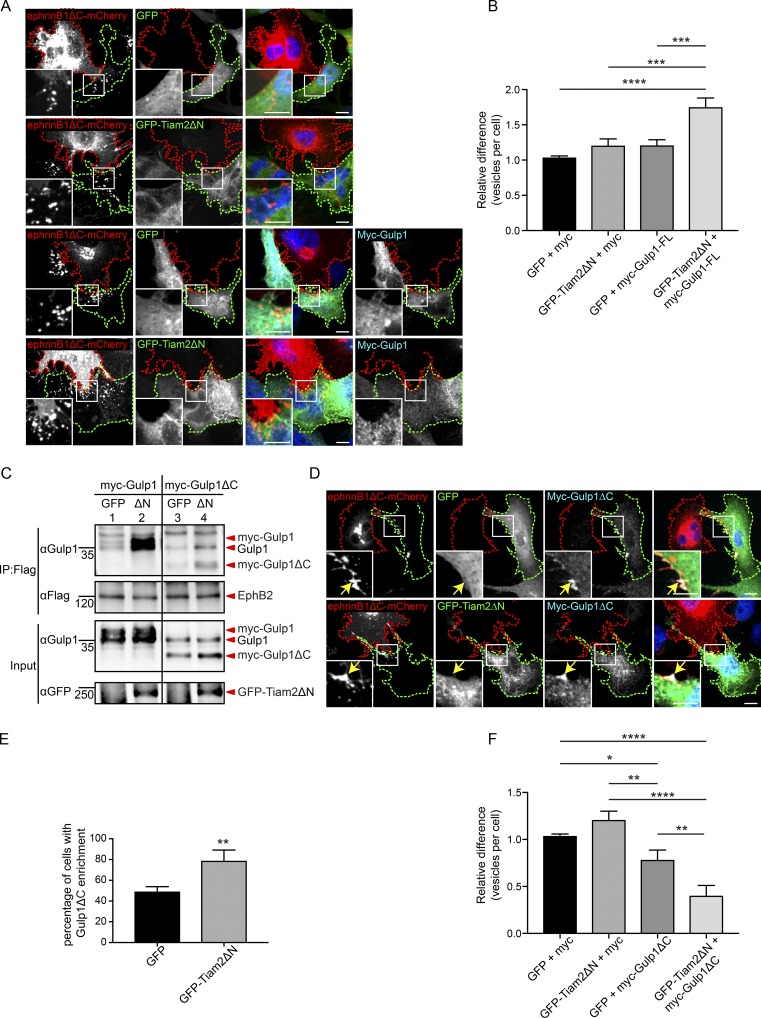
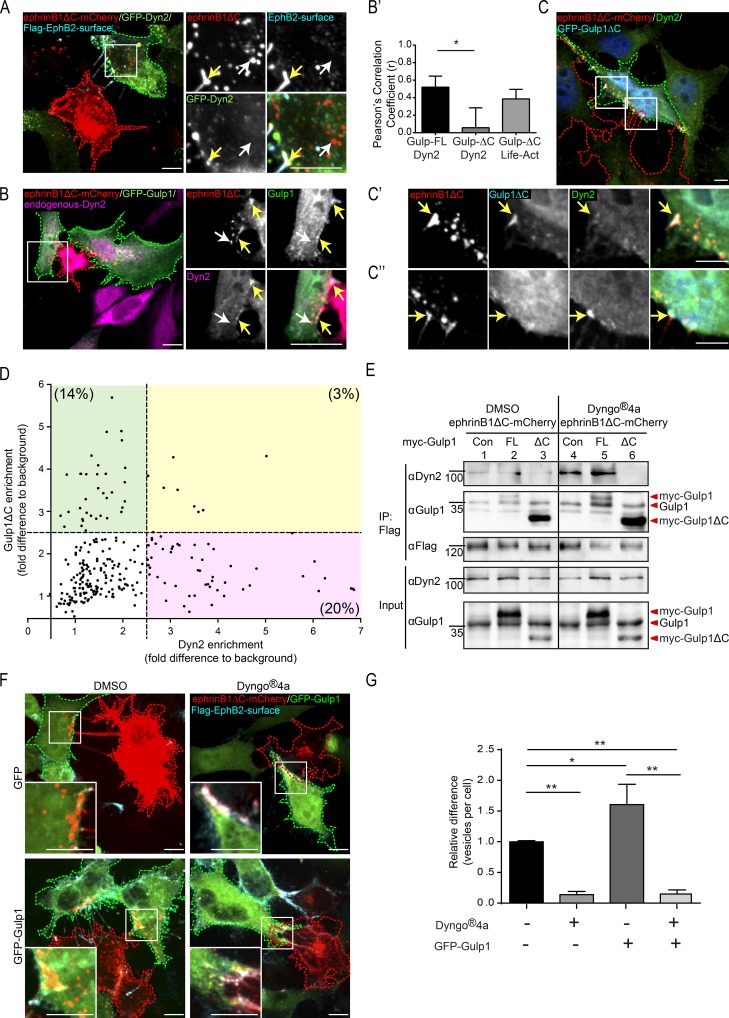
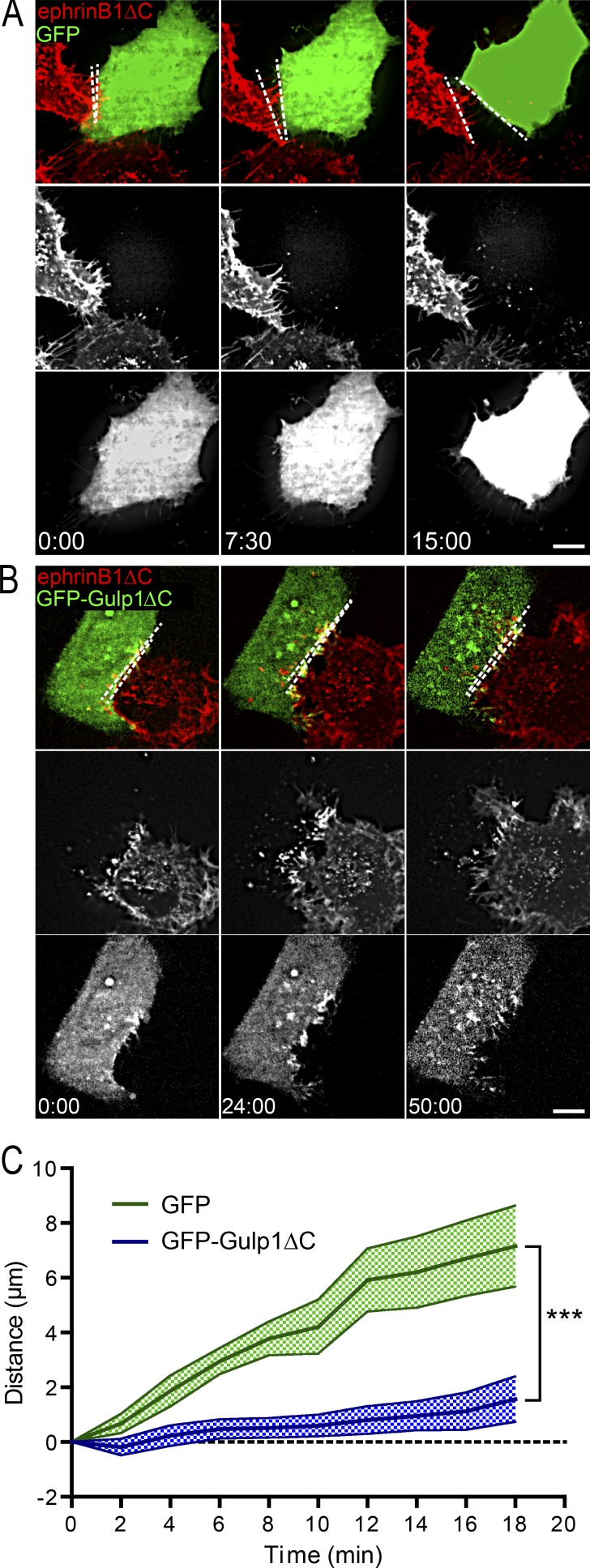
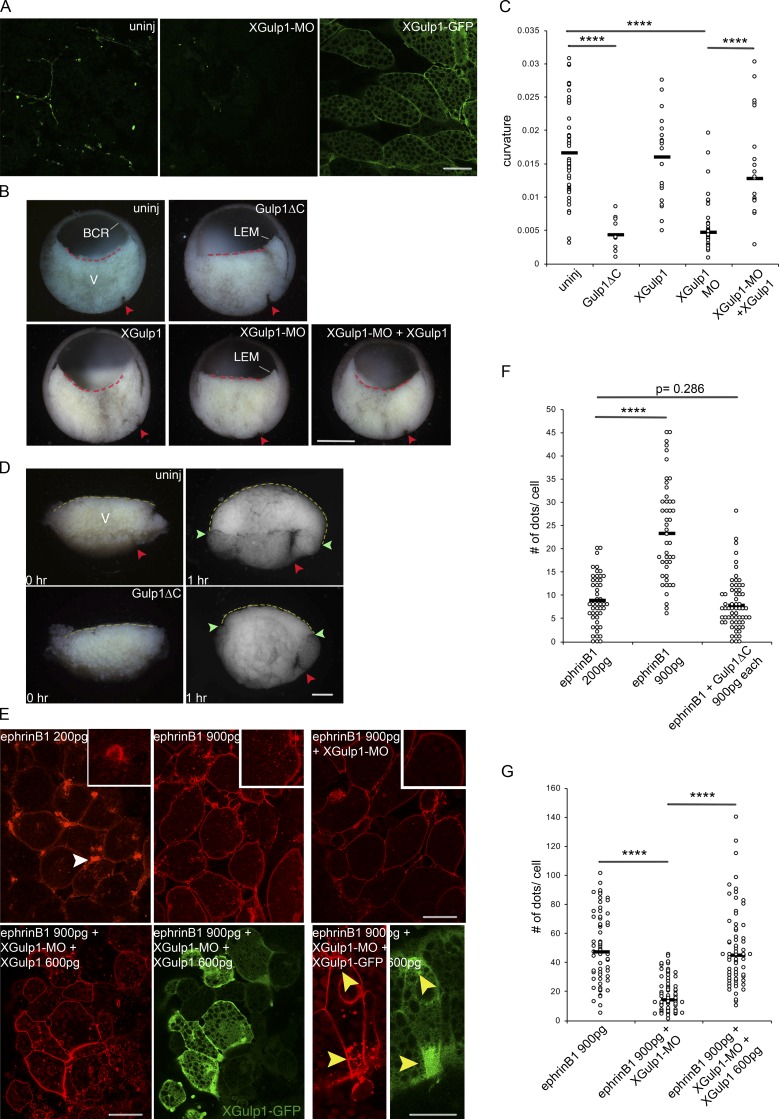
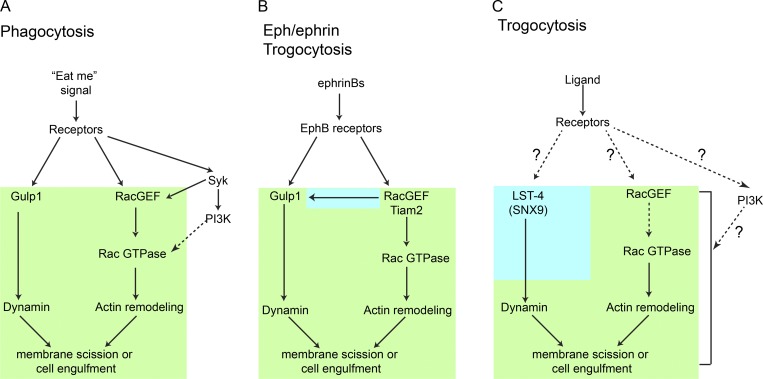
Trogocytosis, in which cells nibble away parts of neighboring cells, is an intercellular cannibalism process conserved from protozoa to mammals. Its underlying molecular mechanisms are not well understood and are likely distinct from phagocytosis, a process that clears entire cells. Bi-directional contact repulsion induced by Eph/ephrin signaling involves transfer of membrane patches and full-length Eph/ephrin protein complexes between opposing cells, resembling trogocytosis. Here, we show that the phagocytic adaptor protein Gulp1 regulates EphB/ephrinB trogocytosis to achieve efficient cell rearrangements of cultured cells and during embryonic development. Gulp1 mediates trogocytosis bi-directionally by dynamic engagement with EphB/ephrinB protein clusters in cooperation with the Rac-specific guanine nucleotide exchange factor Tiam2. Ultimately, Gulp1's presence at the Eph/ephrin cluster is a prerequisite for recruiting the endocytic GTPase dynamin. These results suggest that EphB/ephrinB trogocytosis, unlike other trogocytosis events, uses a phagocytosis-like mechanism to achieve efficient membrane scission and engulfment.
© 2019 Gong et al.
Figures
Comment in
-
Eph receptors and ephrins engage in cellular cannibalism.J Cell Biol. 2019 Oct 7;218(10):3168-3170. doi: 10.1083/jcb.201908074. Epub 2019 Sep 19. J Cell Biol. 2019. PMID: 31537710 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
