

Identification of the Cell-Surface Protease ADAM9 as an Entry Factor for Encephalomyocarditis Virus
- PMID: 31409686
- PMCID: PMC6692517
- DOI: 10.1128/mBio.01780-19
Identification of the Cell-Surface Protease ADAM9 as an Entry Factor for Encephalomyocarditis Virus
Abstract
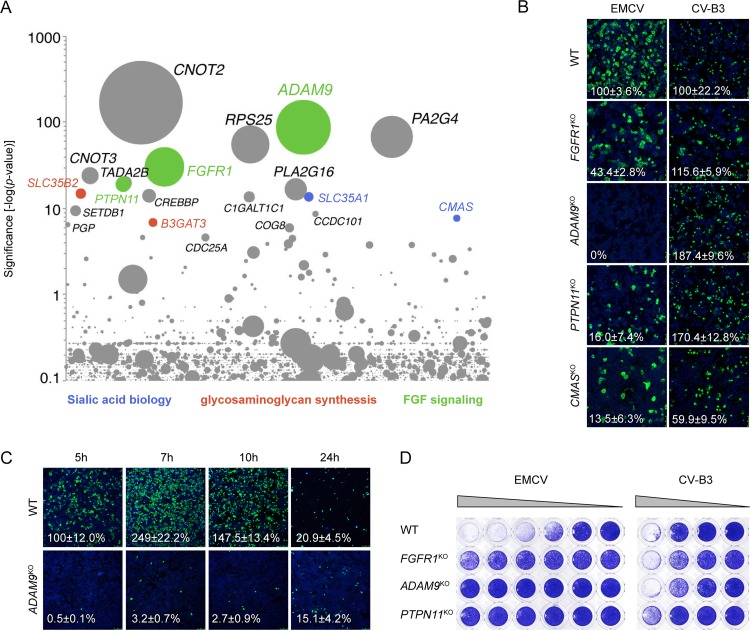
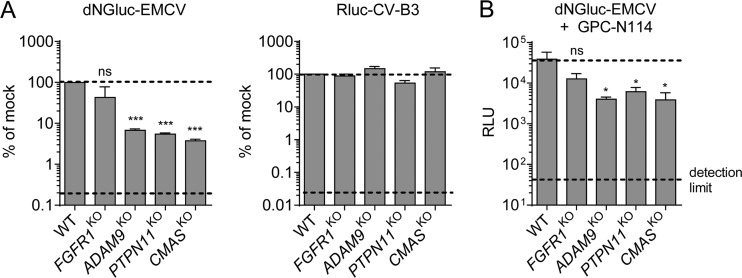
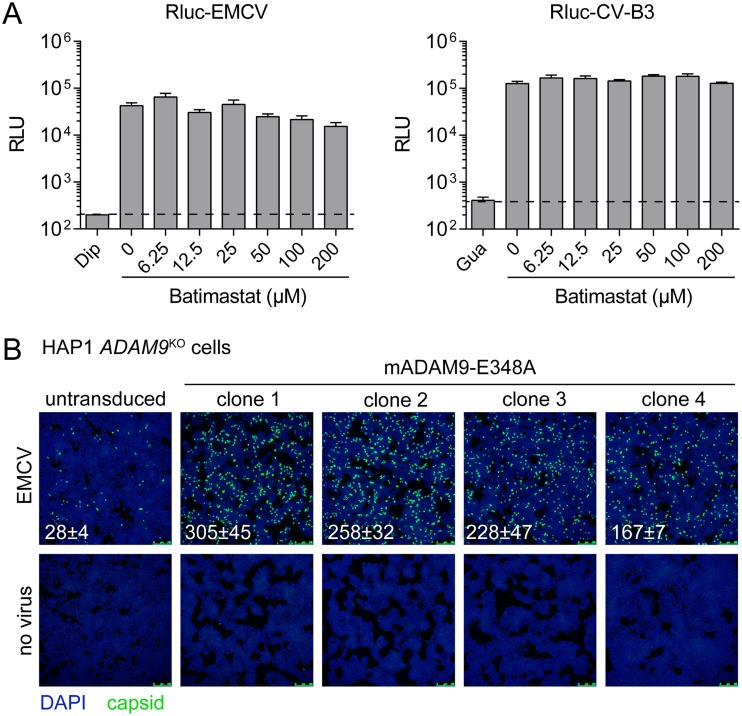
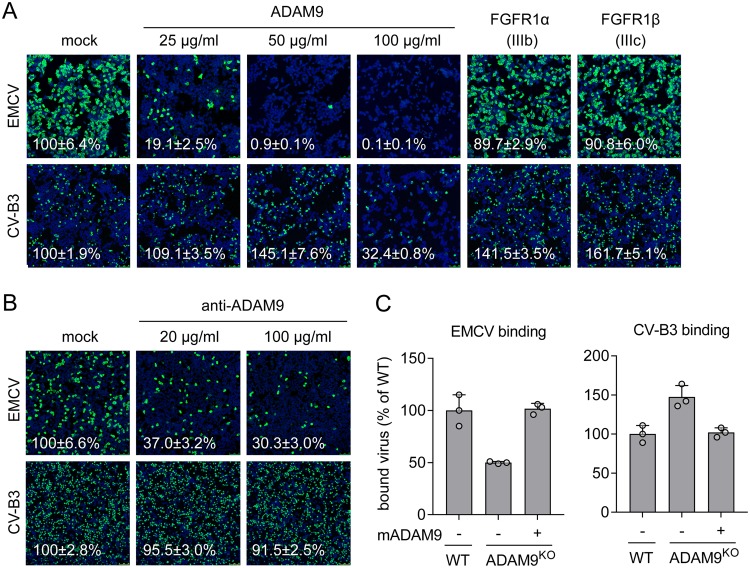
Encephalomyocarditis virus (EMCV) is an animal pathogen and an important model organism, whose receptor requirements are poorly understood. Here, we employed a genome-wide haploid genetic screen to identify novel EMCV host factors. In addition to the previously described picornavirus receptors sialic acid and glycosaminoglycans, this screen unveiled important new host factors for EMCV. These factors include components of the fibroblast growth factor (FGF) signaling pathway, such as the potential receptors FGFR1 and ADAM9, a cell-surface metalloproteinase. By employing various knockout cells, we confirmed the importance of the identified host factors for EMCV infection. The largest reduction in infection efficiency was observed in cells lacking ADAM9. Pharmacological inhibition of the metalloproteinase activity of ADAM9 did not affect virus infection. Moreover, reconstitution of inactive ADAM9 in knockout cells restored susceptibility to EMCV, pointing to a proteinase-independent role of ADAM9 in mediating EMCV infection. Using neutralization assays with ADAM9-specific antiserum and soluble receptor proteins, we provided evidence for a role of ADAM9 in EMCV entry. Finally, binding assays showed that ADAM9 facilitates attachment of EMCV to the cell surface. Together, our findings reveal a role for ADAM9 as a novel receptor or cofactor for EMCV.IMPORTANCE EMCV is an animal pathogen that causes acute viral infections, usually myocarditis or encephalitis. It is thought to circulate mainly among rodents, from which it is occasionally transmitted to other animal species, including humans. EMCV causes fatal outbreaks of myocarditis and encephalitis in pig farms and zoos, making it an important veterinary pathogen. Although EMCV has been widely used as a model to study mechanisms of viral disease in mice, little is known about its entry mechanism. Here, we employ a haploid genetic screen for EMCV host factors and identify an essential role for ADAM9 in EMCV entry.
Keywords: disintegrin and metalloproteinase domain-containing protein 9 (ADAM9); encephalomyocarditis virus; haploid genetic screen.
Copyright © 2019 Baggen et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
