

AtNDB2 Is the Main External NADH Dehydrogenase in Mitochondria and Is Important for Tolerance to Environmental Stress
- PMID: 31409698
- PMCID: PMC6776847
- DOI: 10.1104/pp.19.00877
AtNDB2 Is the Main External NADH Dehydrogenase in Mitochondria and Is Important for Tolerance to Environmental Stress
Abstract
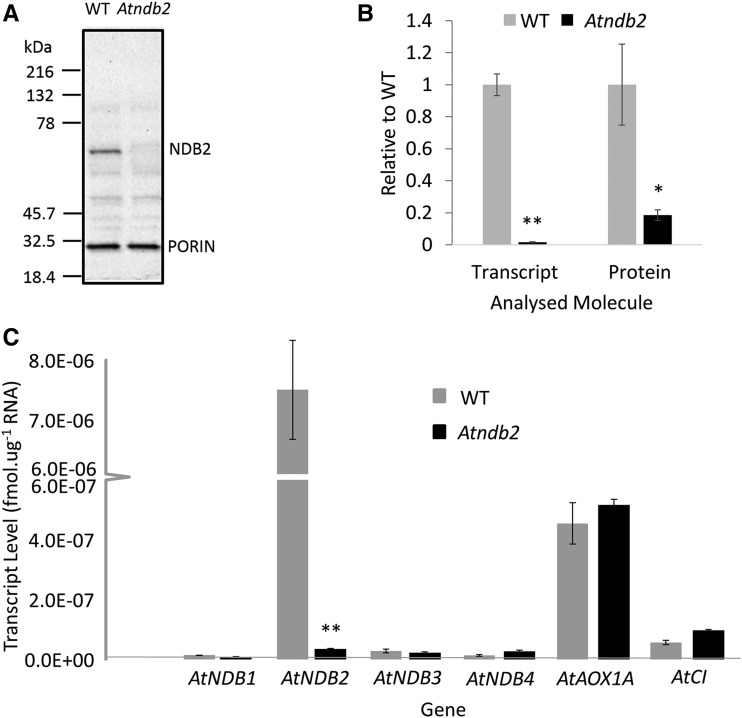
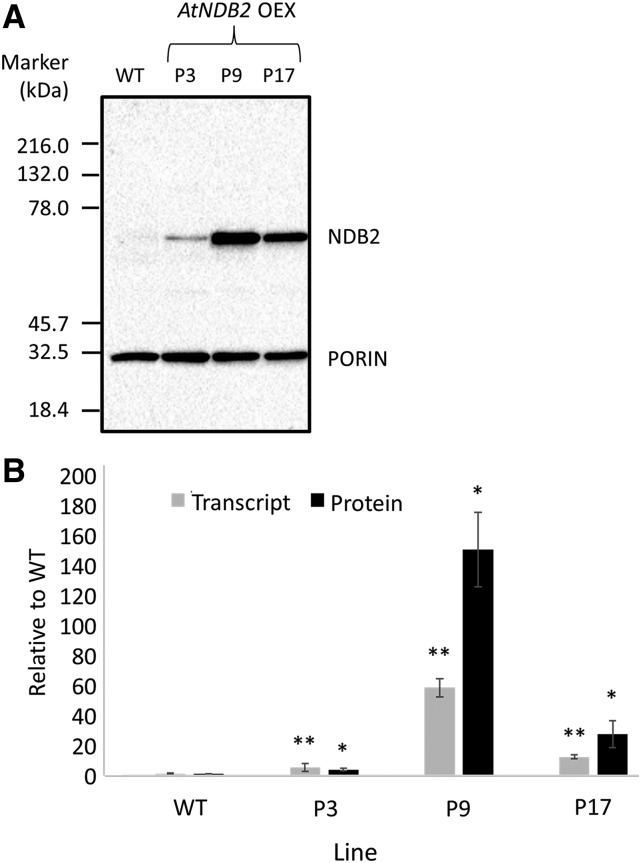
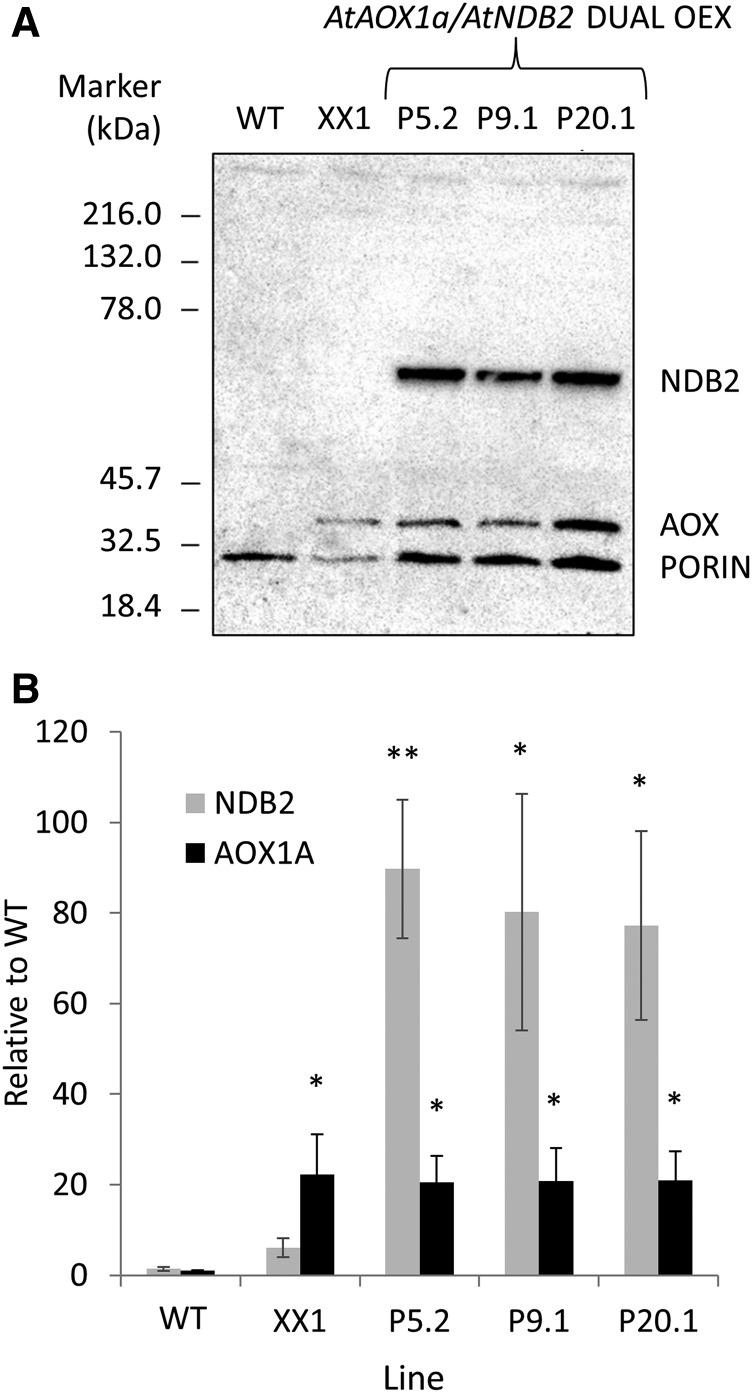
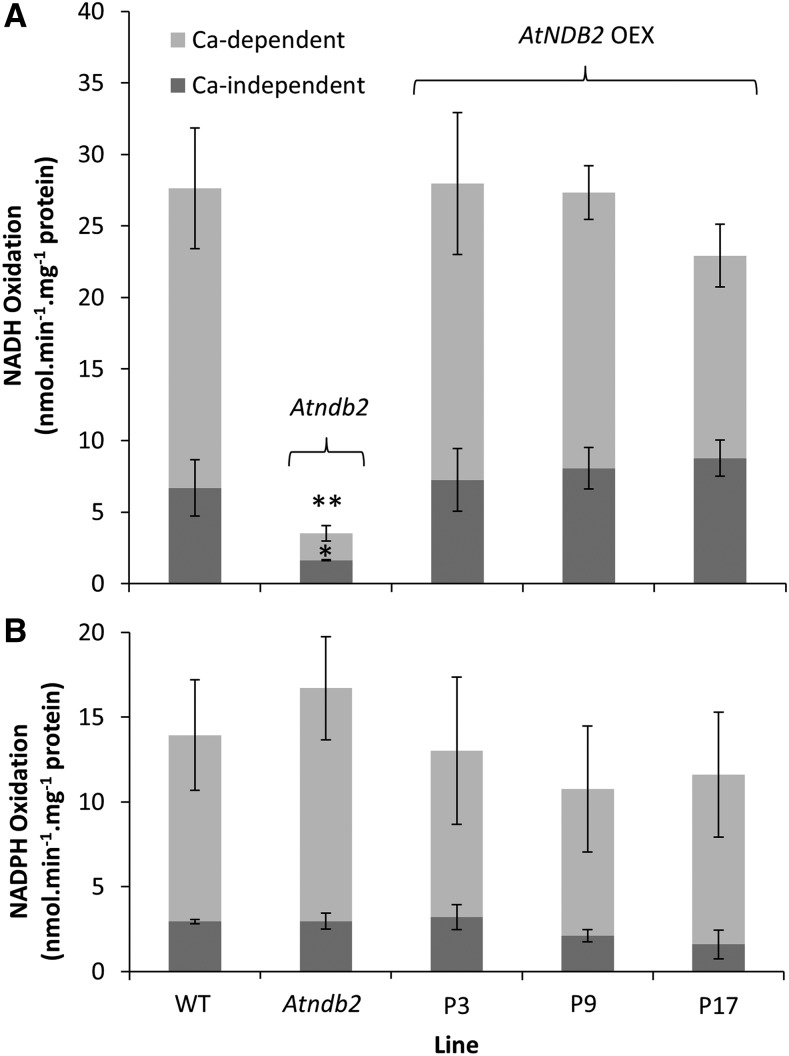
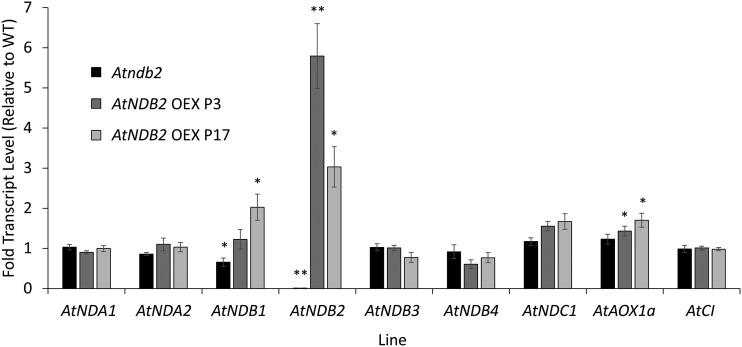
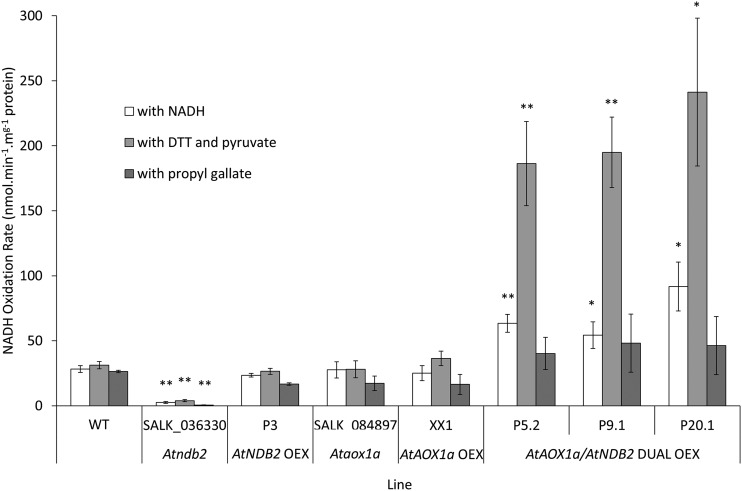
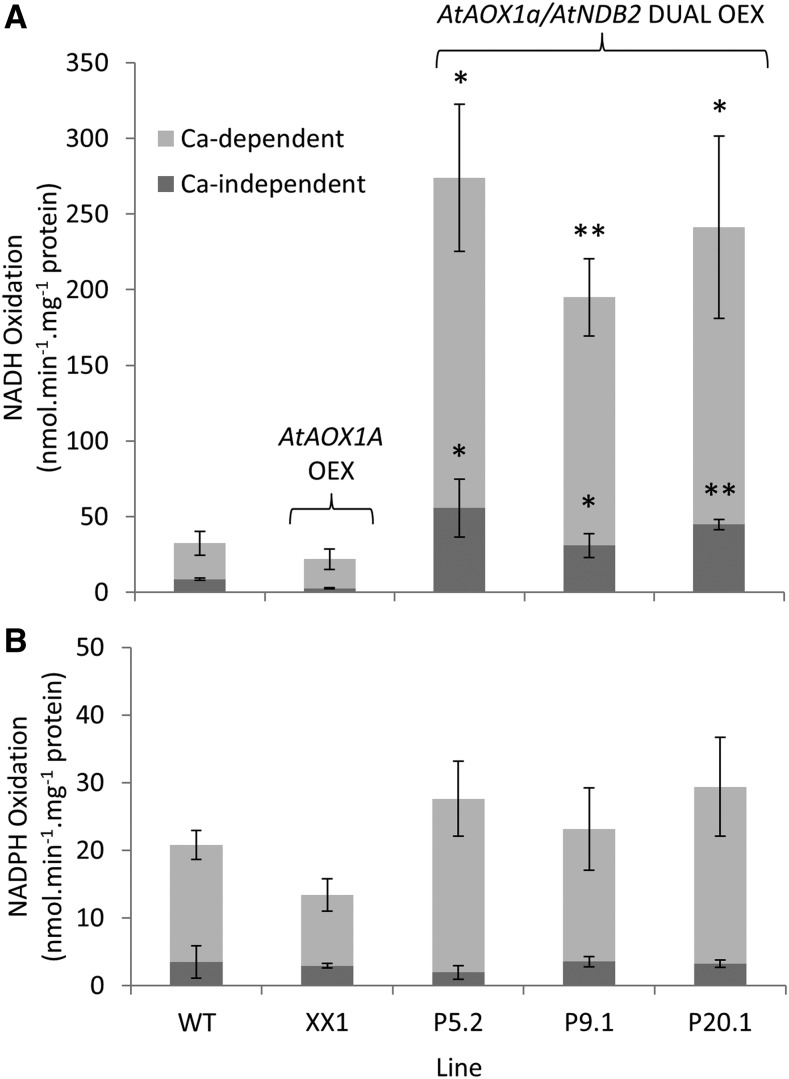
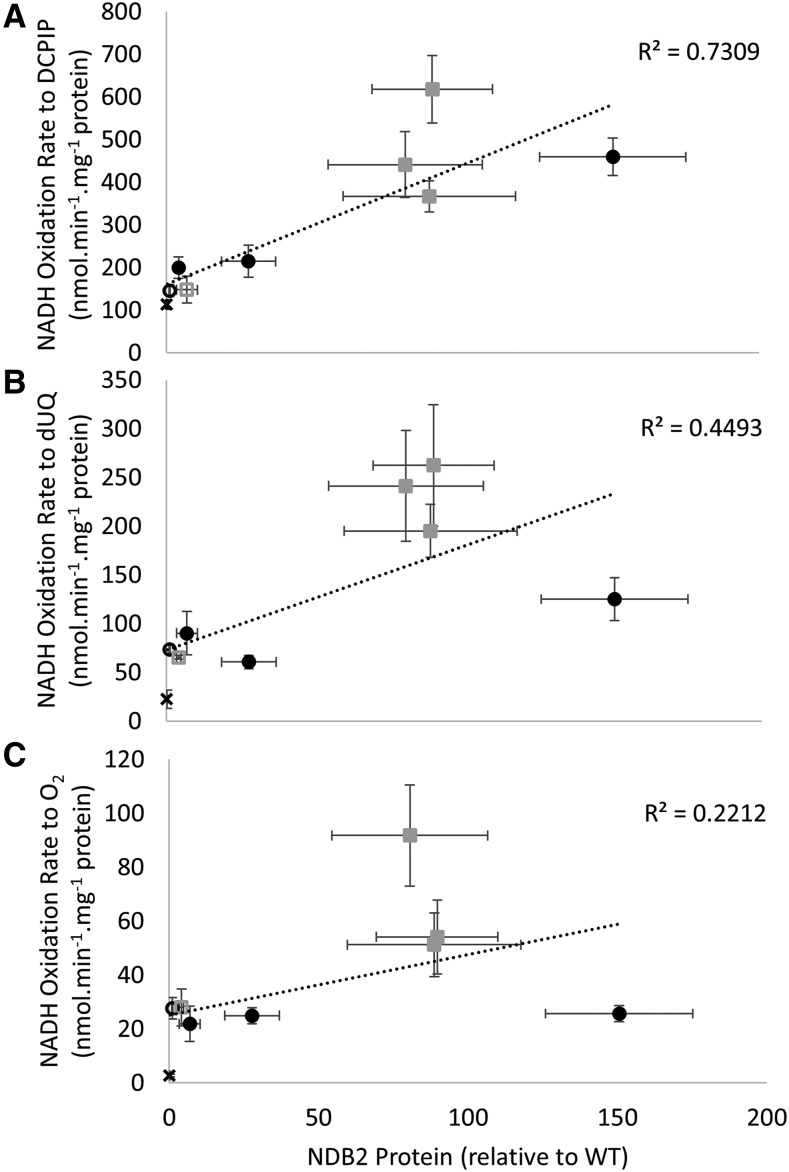
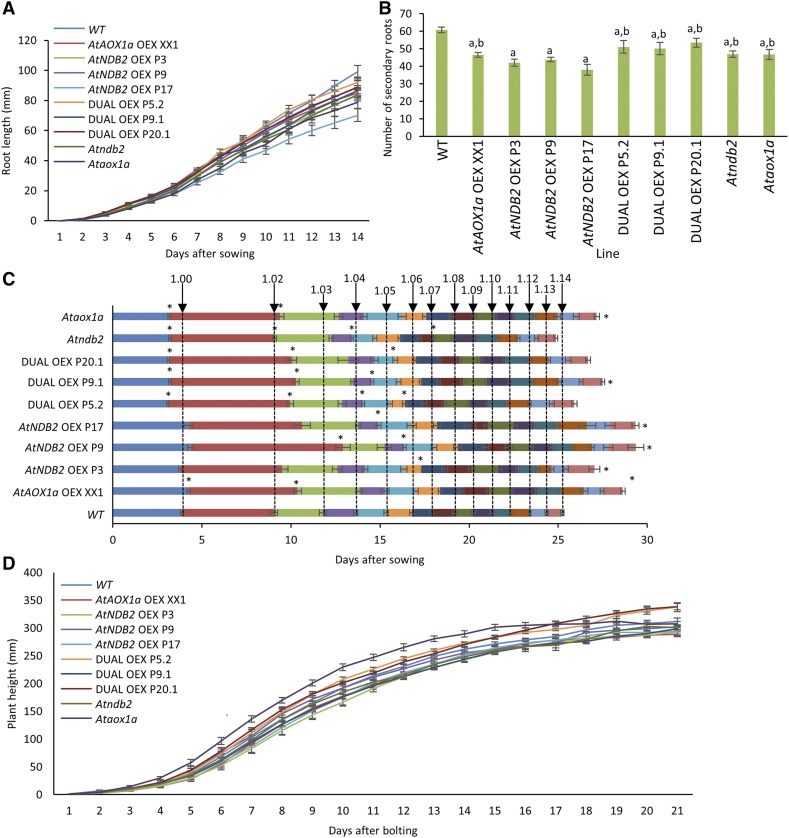
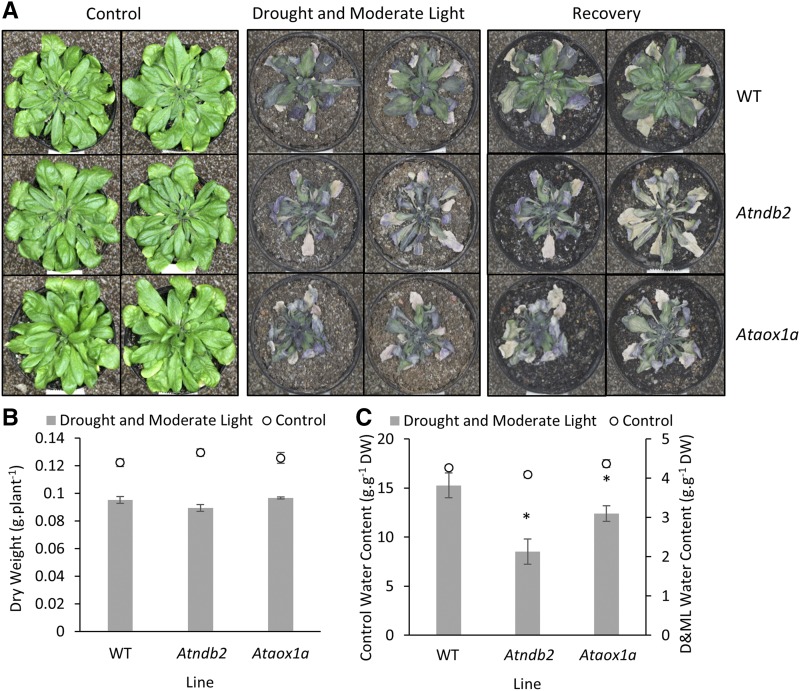
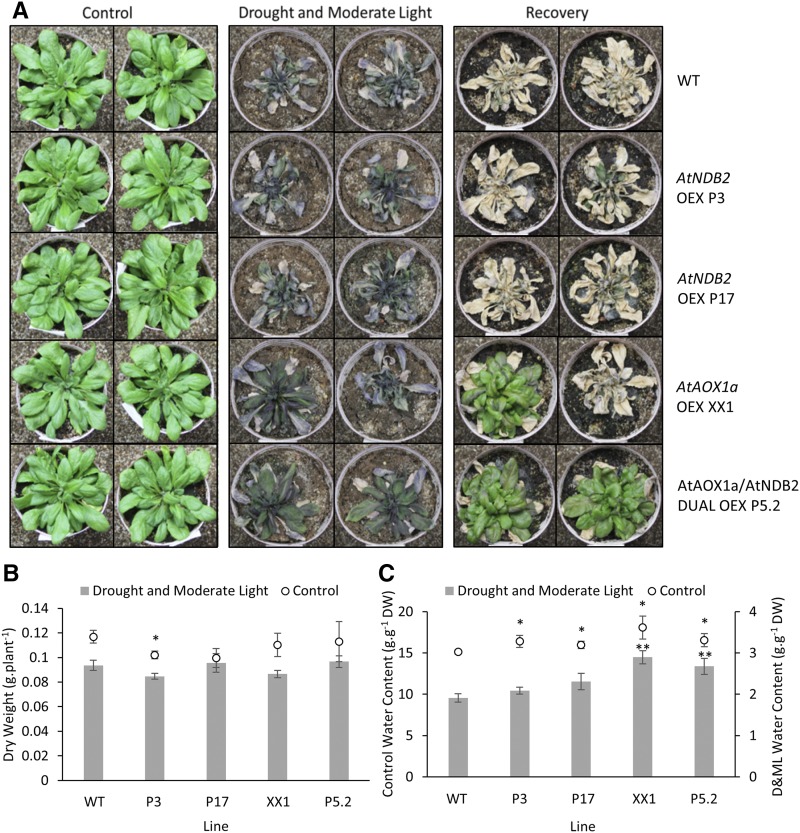
In addition to the classical electron transport pathway coupled to ATP synthesis, plant mitochondria have an alternative pathway that involves type II NAD(P)H dehydrogenases (NDs) and alternative oxidase (AOX). This alternative pathway participates in thermogenesis in select organs of some species and is thought to help prevent cellular damage during exposure to environmental stress. Here, we investigated the function and role of one alternative path component, AtNDB2, using a transgenic approach in Arabidopsis (Arabidopsis thaliana). Disruption of AtNDB2 expression via T-DNA insertion led to a 90% decrease of external NADH oxidation in isolated mitochondria. Overexpression of AtNDB2 led to increased AtNDB2 protein abundance in mitochondria but did not enhance external NADH oxidation significantly unless AtAOX1A was concomitantly overexpressed and activated, demonstrating a functional link between these enzymes. Plants lacking either AtAOX1A or AtNDB2 were more sensitive to combined drought and elevated light treatments, whereas plants overexpressing these components showed increased tolerance and capacity for poststress recovery. We conclude that AtNDB2 is the predominant external NADH dehydrogenase in mitochondria and together with AtAOX1A forms a complete, functional, nonphosphorylating pathway of electron transport, whose operation enhances tolerance to environmental stress. This study demonstrates that at least one of the alternative NDs, as well as AOX, are important for the stress response.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Armstrong AF, Badger MR, Day DA, Barthet MM, Smith PMC, Millar AH, Whelan J, Atkin OK (2008) Dynamic changes in the mitochondrial electron transport chain underpinning cold acclimation of leaf respiration. Plant Cell Environ 31: 1156–1169 - PubMed
-
- Arnholdt-Schmitt B, Costa JH, de Melo DF (2006) AOX: A functional marker for efficient cell reprogramming under stress? Trends Plant Sci 11: 281–287 - PubMed
-
- Bartoli CG, Yu J, Gómez F, Fernández L, McIntosh L, Foyer CH (2006) Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J Exp Bot 57: 1621–1631 - PubMed
-
- Carrie C, Murcha MW, Kuehn K, Duncan O, Barthet M, Smith PM, Eubel H, Meyer E, Day DA, Millar AH, et al. (2008) Type II NAD(P)H dehydrogenases are targeted to mitochondria and chloroplasts or peroxisomes in Arabidopsis thaliana. FEBS Lett 582: 3073–3079 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
