

Estrogen receptor β inhibits breast cancer cells migration and invasion through CLDN6-mediated autophagy
- PMID: 31412908
- PMCID: PMC6694553
- DOI: 10.1186/s13046-019-1359-9
Estrogen receptor β inhibits breast cancer cells migration and invasion through CLDN6-mediated autophagy
Abstract
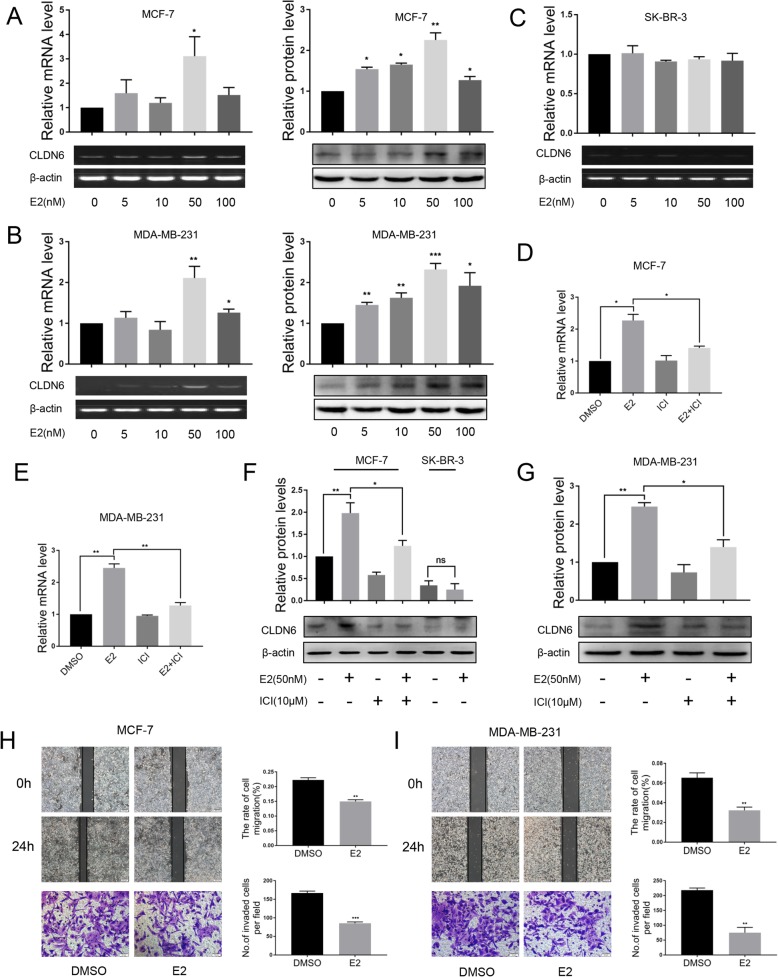
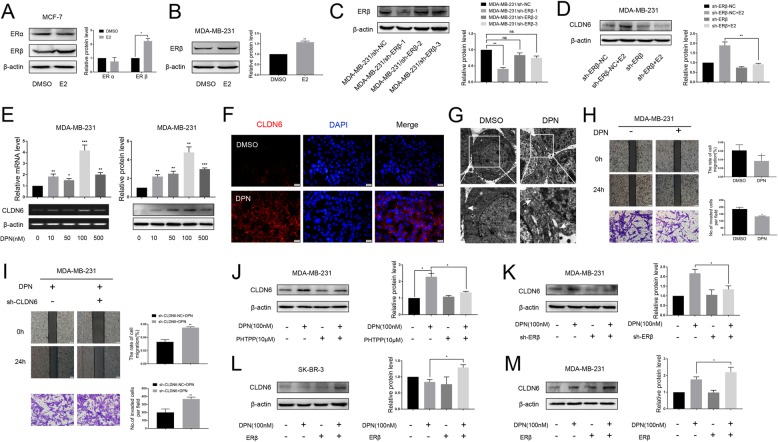
Background: Estrogen receptor β (ERβ) has been reported to play an anti-cancer role in breast cancer, but the regulatory mechanism by which ERβ exerts this effect is not clear. Claudin-6 (CLDN6), a tight junction protein, acts as a tumor suppressor gene in breast cancer. Our previous studies have found that 17β-estradiol (E2) induces CLDN6 expression and inhibits MCF-7 cell migration and invasion, but the underlying molecular mechanisms are still unclear. In this study, we aimed to investigate the role of ERβ in this process and the regulatory mechanisms involved.
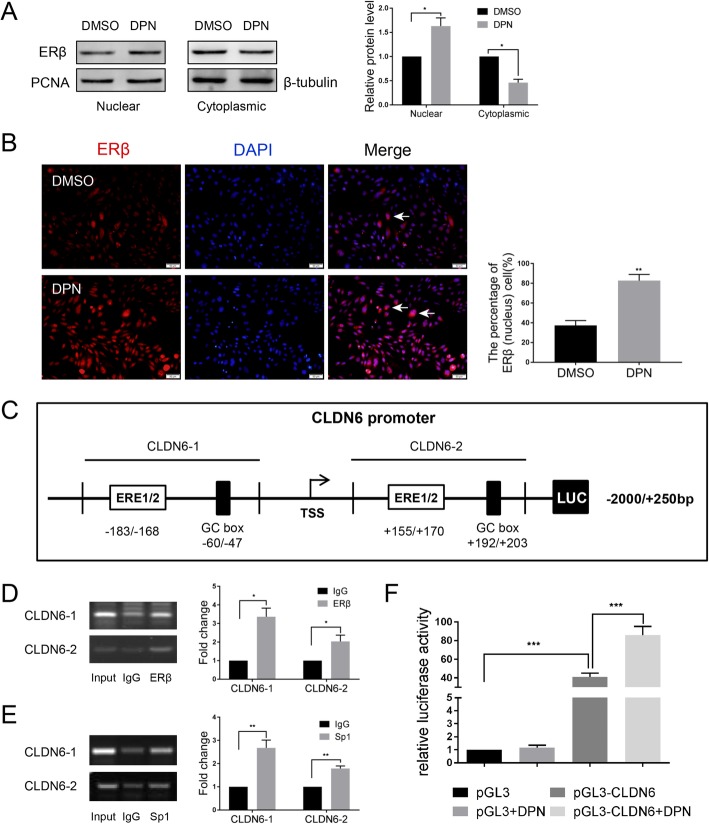
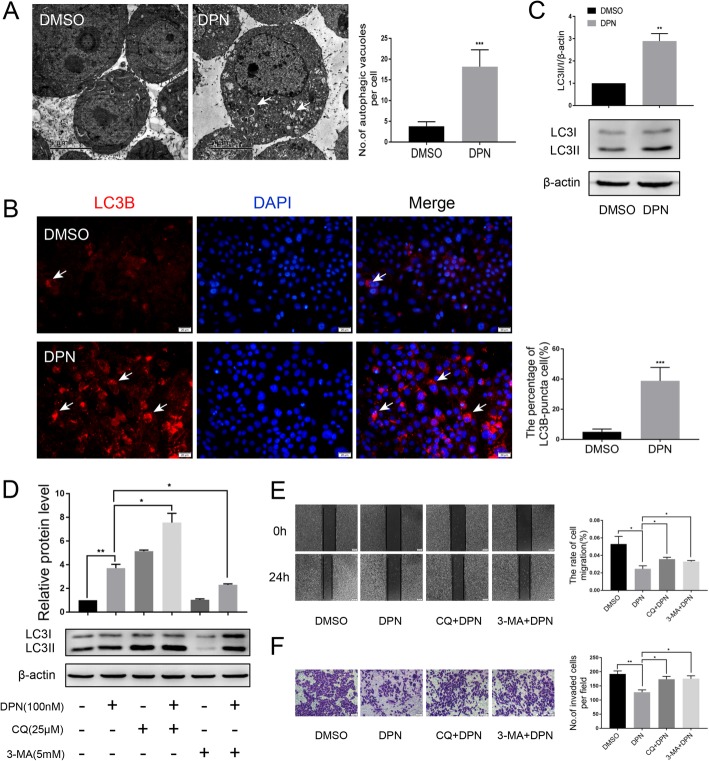
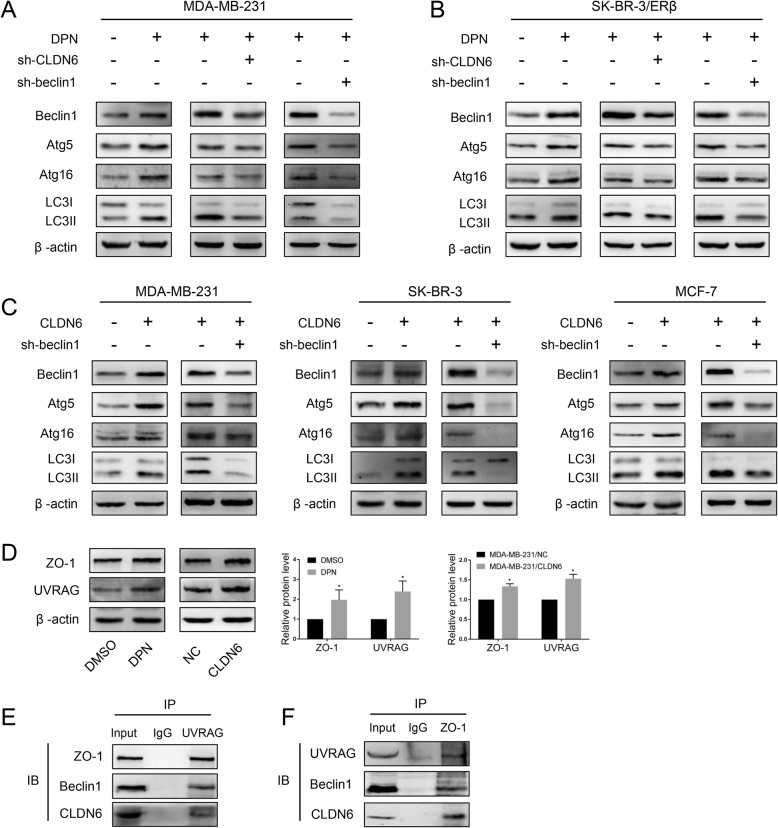
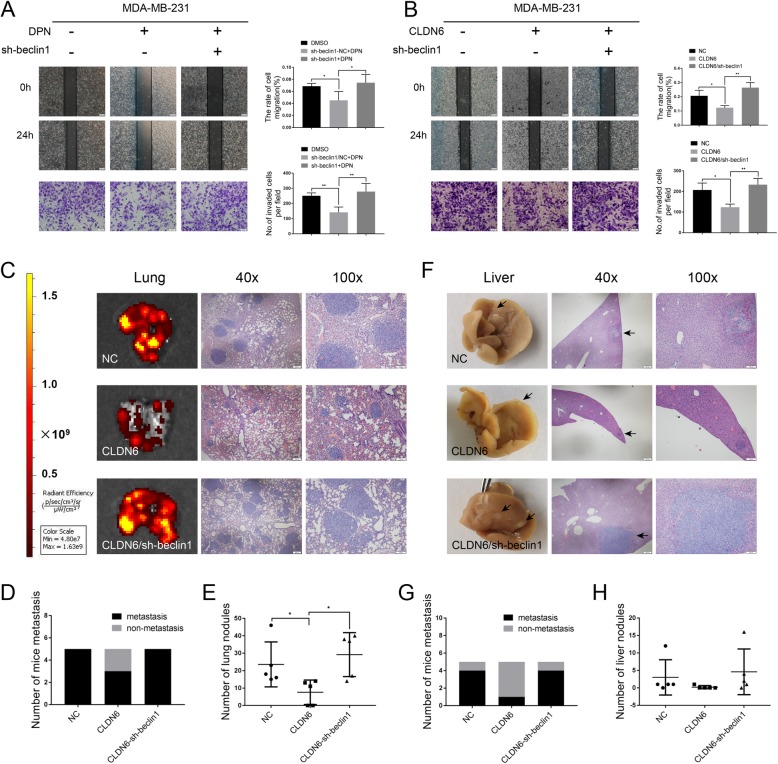
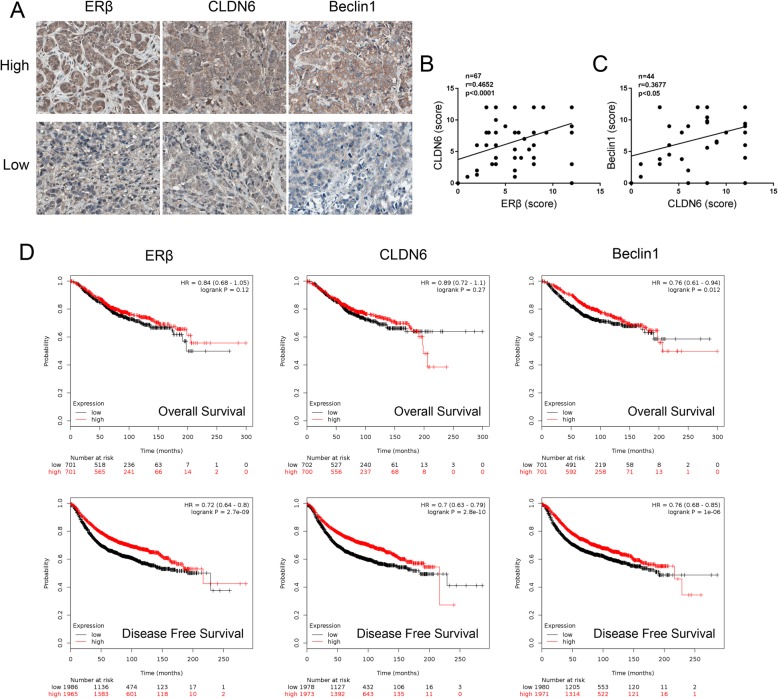
Methods: Polymerase chain reaction (PCR) and western blot were used to characterize the effect of E2 on the expression of CLDN6 in breast cancer cells. Chromatin immunoprecipitation (ChIP) assays were carried out to confirm the interaction between ERβ and CLDN6. Dual luciferase reporter assays were used to detect the regulatory role of ERβ on the promoter activity of CLDN6. Wound healing and Transwell assays were used to examine the migration and invasion of breast cancer cells. Western blot, immunofluorescence and transmission electron microscopy (TEM) were performed to detect autophagy. Xenograft mouse models were used to explore the regulatory effect of the CLDN6-beclin1 axis on breast cancer metastasis. Immunohistochemistry (IHC) was used to detect ERβ/CLDN6/beclin1 expression in breast cancer patient samples.
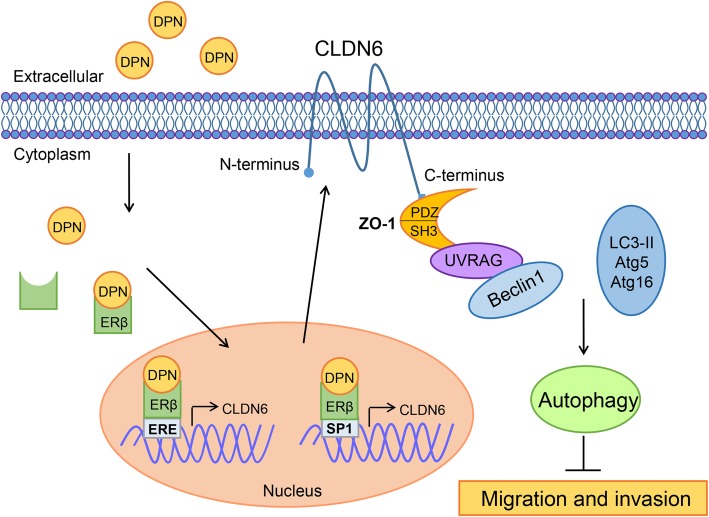
Results: Here, E2 upregulated the expression of CLDN6, which was mediated by ERβ. ERβ regulated CLDN6 expression at the transcriptional level. ERβ inhibited the migration and invasion of breast cancer cells through CLDN6. Interestingly, this effect was associated with CLDN6-induced autophagy. CLDN6 positively regulated the expression of beclin1, which is a key regulator of autophagy. Beclin1 knockdown reversed CLDN6-induced autophagy and the inhibitory effect of CLDN6 on breast cancer metastasis. Moreover, ERβ and CLDN6 were positively correlated, and the expression of CLDN6 was positively correlated with beclin1 in breast cancer tissues.
Conclusion: Overall, this is the first study to demonstrate that the inhibitory effect of ERβ on the migration and invasion of breast cancer cells was mediated by CLDN6, which induced the beclin1-dependent autophagic cascade.
Keywords: Autophagy; Breast cancer; CLDN6; Estrogen receptor β; Invasion; Migration.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
