

Dengue Virus M Protein Promotes NLRP3 Inflammasome Activation To Induce Vascular Leakage in Mice
- PMID: 31413130
- PMCID: PMC6803285
- DOI: 10.1128/JVI.00996-19
Dengue Virus M Protein Promotes NLRP3 Inflammasome Activation To Induce Vascular Leakage in Mice
Abstract
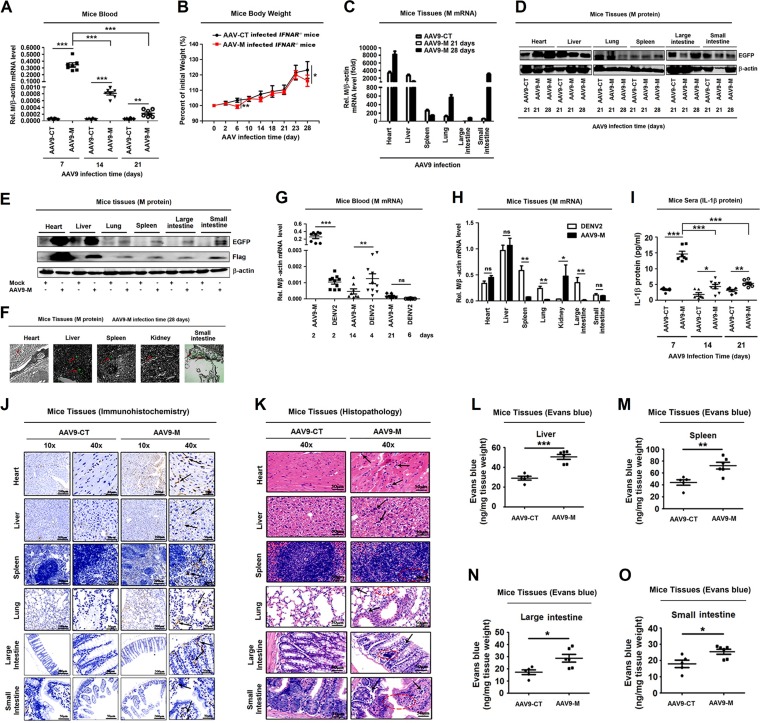
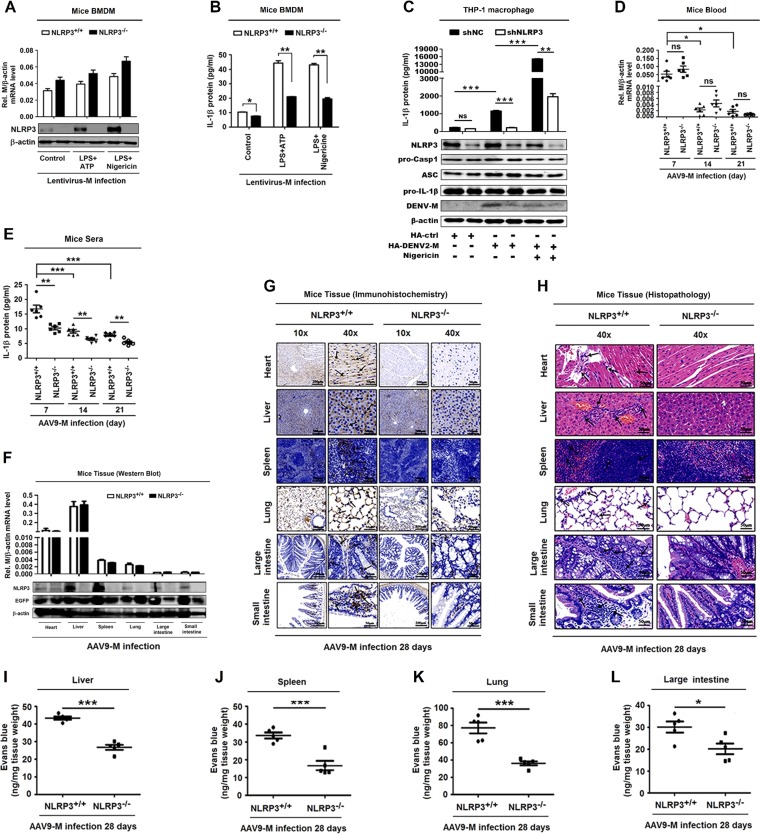
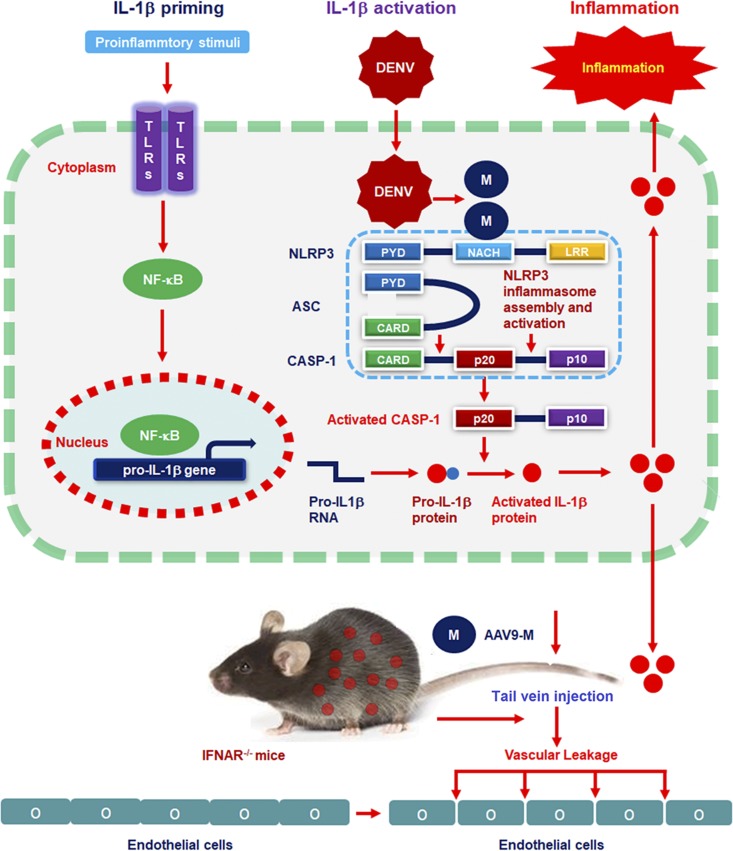
Dengue virus (DENV) infection causes serious clinical symptoms, including dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS). Vascular permeability change is the main feature of the diseases, and the abnormal expression of proinflammatory cytokines is the important cause of vascular permeability change. However, the mechanism underlying vascular permeability induced by DENV has not been fully elucidated. Here, we reveal a distinct mechanism by which DENV infection promotes NLRP3 inflammasome activation and interleukin-1 beta (IL-1β) release to induce endothelial permeability and vascular leakage in mice. DENV M protein interacts with NLRP3 to facilitate NLRP3 inflammasome assembly and activation, which induce proinflammatory cytokine IL-1β activation and release. Notably, M can induce vascular leakage in mouse tissues by activating the NLRP3 inflammasome and IL-1β. More importantly, inflammatory cell infiltration and tissue injuries are induced by M in wild-type (WT) mouse tissues, but they are not affected by M in NLRP3 knockout (NLRP3-/-) mouse tissues. Evans blue intensities in WT mouse tissues are significantly higher than in NLRP3-/- mouse tissues, demonstrating an essential role of NLRP3 in M-induced vascular leakages in mice. Therefore, we propose that upon DENV infection, M interacts with NLRP3 to facilitate inflammasome activation and IL-1β secretion, which lead to the induction of endothelial permeability and vascular leakage in mouse tissues. The important role of the DENV-M-NLRP3-IL-1β axis in the induction of vascular leakage provides new insights into the mechanisms underlying DENV pathogenesis and DENV-associated DHF and DSS development.IMPORTANCE Dengue virus (DENV) is a mosquito-borne pathogen, and infections by this virus are prevalent in over 100 tropical and subtropical countries or regions, with approximately 2.5 billion people at risk. DENV infection induces a spectrum of clinical symptoms, ranging from classical dengue fever (DF) to severe dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS). Therefore, it is important to understand the mechanisms underlying DENV pathogenesis. In this study, we reveal that the DENV membrane protein (M) interacts with the host NLRP3 protein to promote NLRP3 inflammasome activation, which leads to the activation and release of a proinflammatory cytokine, interleukin-1 beta (IL-1β). More importantly, we demonstrate that M protein can induce vascular permeability and vascular leakage and that NLRP3 is required for M-induced vascular leakage in mouse tissues. Collectively, this study reveals a distinct mechanism underlying DENV pathogeneses and provides new insights into the development of therapeutic agents for DENV-associated diseases.
Keywords: DENV; DENV membrane protein; IL-1β; M; NLRP3; NLRP3 inflammasome; Nod-like receptor (NLR) family pyrin domain containing 3; dengue virus; interleukin-1β; vascular leakage; vascular permeability.
Copyright © 2019 American Society for Microbiology.
Figures
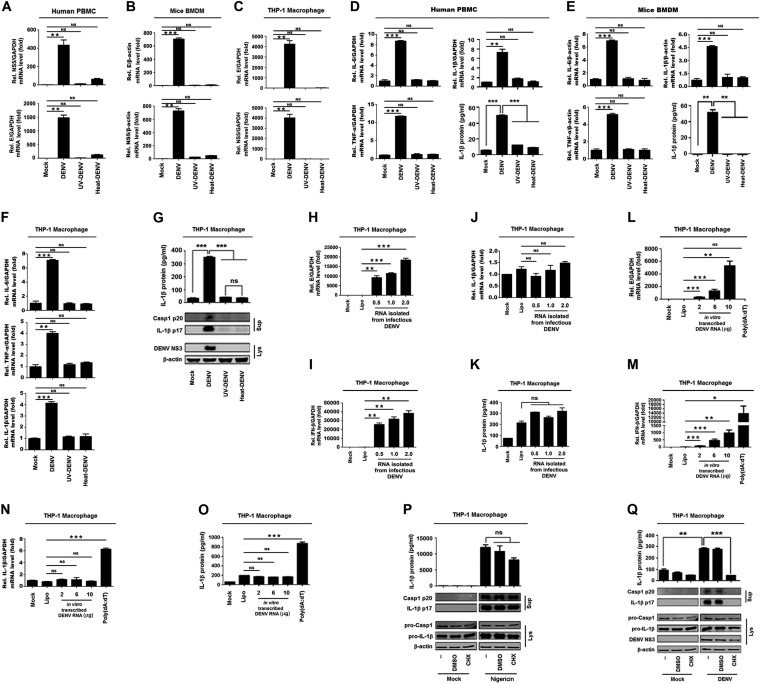
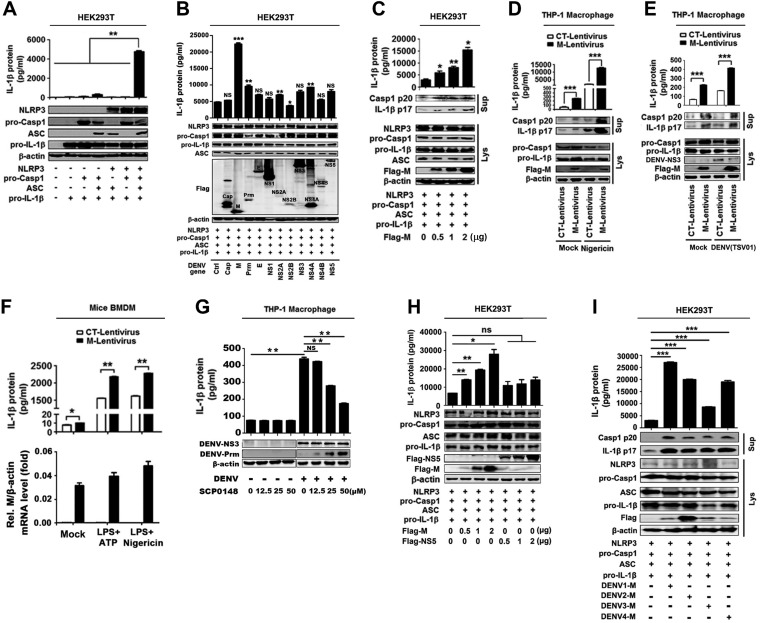
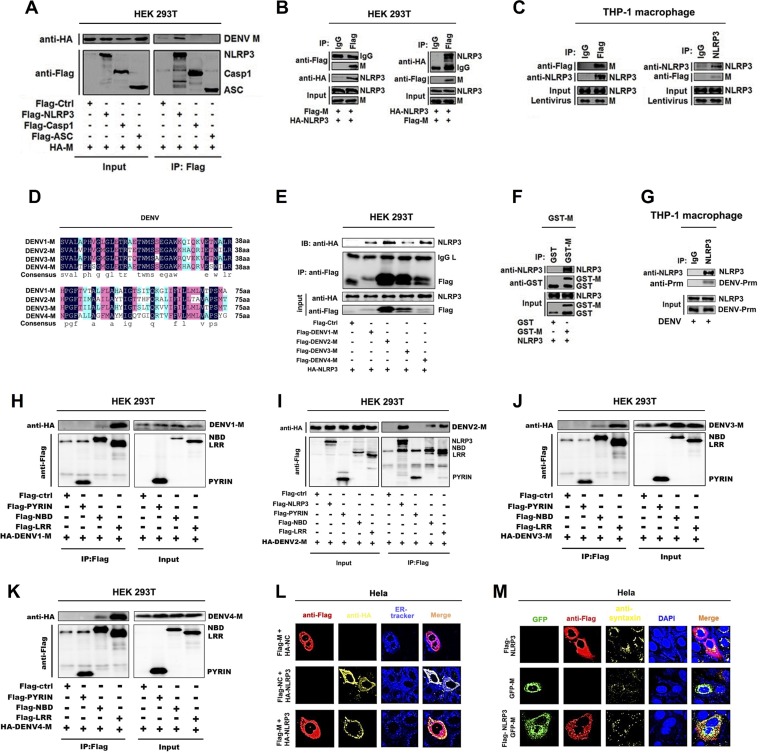
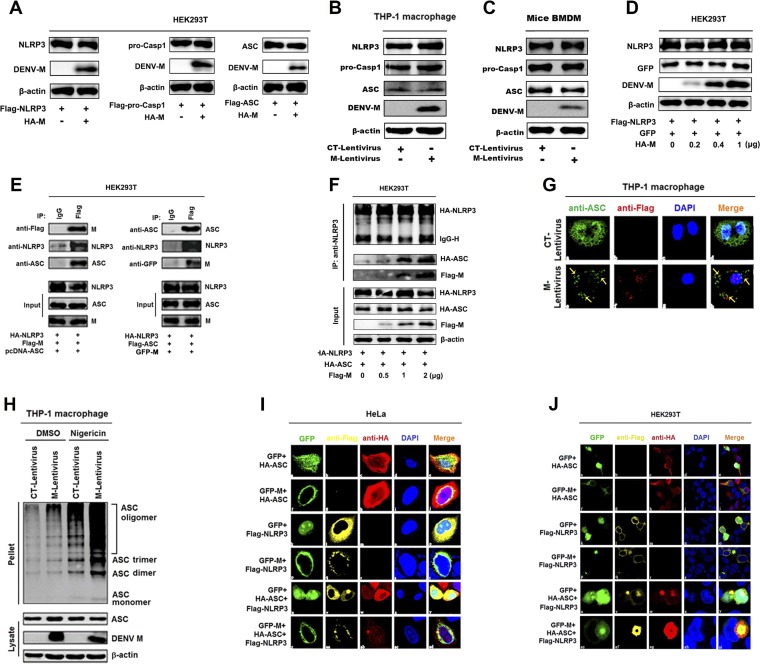
References
-
- Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. 2013. The global distribution and burden of dengue. Nature 496:504–507. doi:10.1038/nature12060. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
