

Toxoplasma gondii effector TgIST blocks type I interferon signaling to promote infection
- PMID: 31413201
- PMCID: PMC6717281
- DOI: 10.1073/pnas.1904637116
Toxoplasma gondii effector TgIST blocks type I interferon signaling to promote infection
Abstract
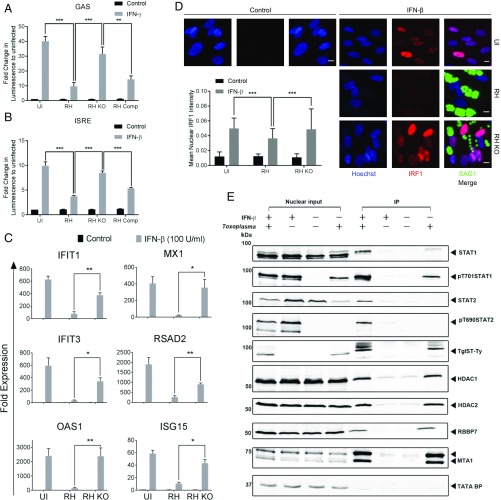
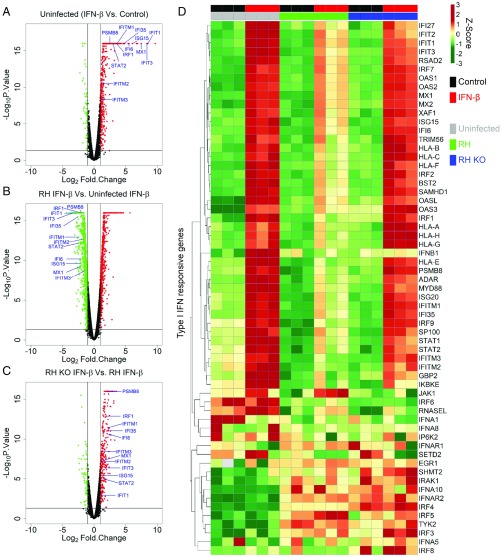
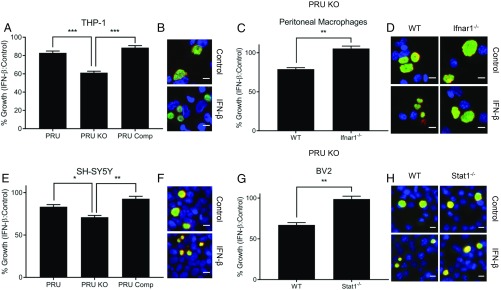
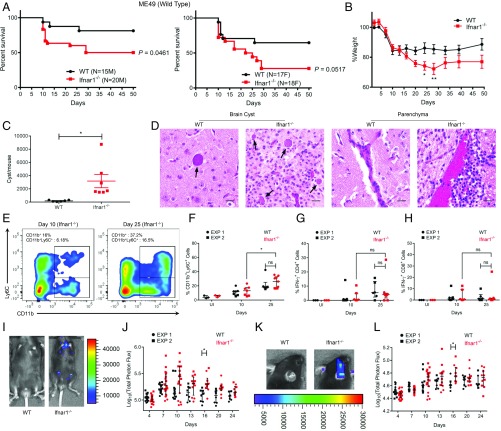
In contrast to the importance of type II interferon-γ (IFN-γ) in control of toxoplasmosis, the role of type I IFN is less clear. We demonstrate here that TgIST, a secreted effector previously implicated in blocking type II IFN-γ signaling, also blocked IFN-β responses by inhibiting STAT1/STAT2-mediated transcription in infected cells. Consistent with a role for type I IFN in cell intrinsic control, ∆Tgist mutants were more susceptible to growth inhibition by murine and human macrophages activated with IFN-β. Additionally, type I IFN was important for production of IFN-γ by natural killer (NK) cells and recruitment of inflammatory monocytes at the site of infection. Mice lacking type I IFN receptors (Ifnar1-/-) showed increased mortality following infection with wild-type parasites and decreased virulence of ∆Tgist parasites was restored in Ifnar1-/- mice. The findings highlight the importance of type I IFN in control of toxoplasmosis and illuminate a parasite mechanism to counteract the effects of both type I and II IFN-mediated host defenses.
Keywords: NK cell; central nervous system; inflammatory monocyte; interferon; transcriptome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Reich N. C., Liu L., Tracking STAT nuclear traffic. Nat. Rev. Immunol. 6, 602–612 (2006). - PubMed
-
- Schroder K., Hertzog P. J., Ravasi T., Hume D. A., Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 75, 163–189 (2004). - PubMed
-
- Ramana C. V., Chatterjee-Kishore M., Nguyen H., Stark G. R., Complex roles of Stat1 in regulating gene expression. Oncogene 19, 2619–2627 (2000). - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
