

Insights into ubiquitin chain architecture using Ub-clipping
- PMID: 31413367
- PMCID: PMC6823057
- DOI: 10.1038/s41586-019-1482-y
Insights into ubiquitin chain architecture using Ub-clipping
Abstract
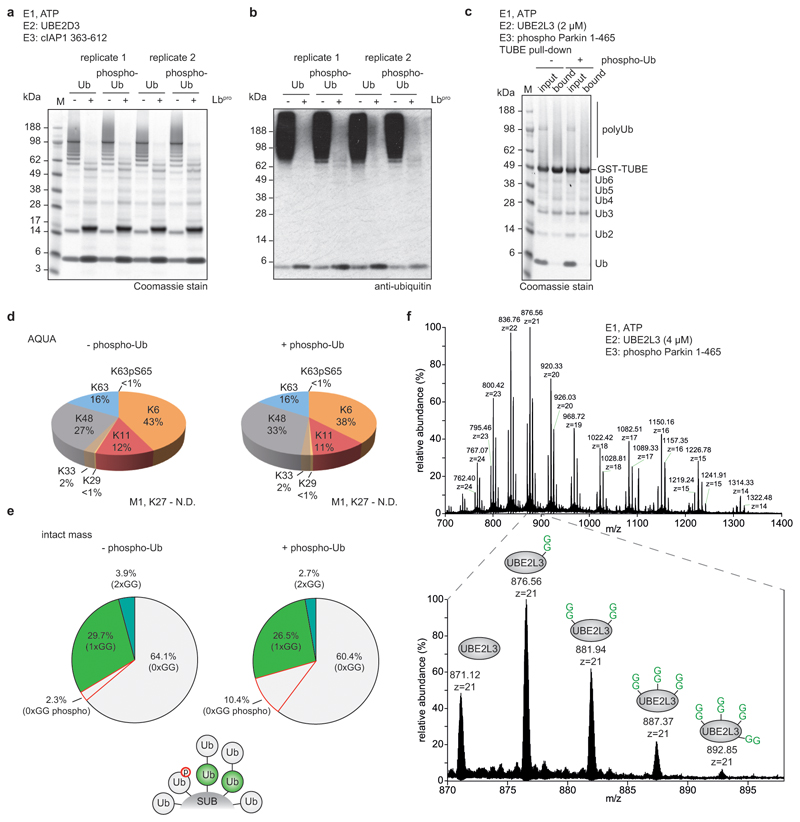
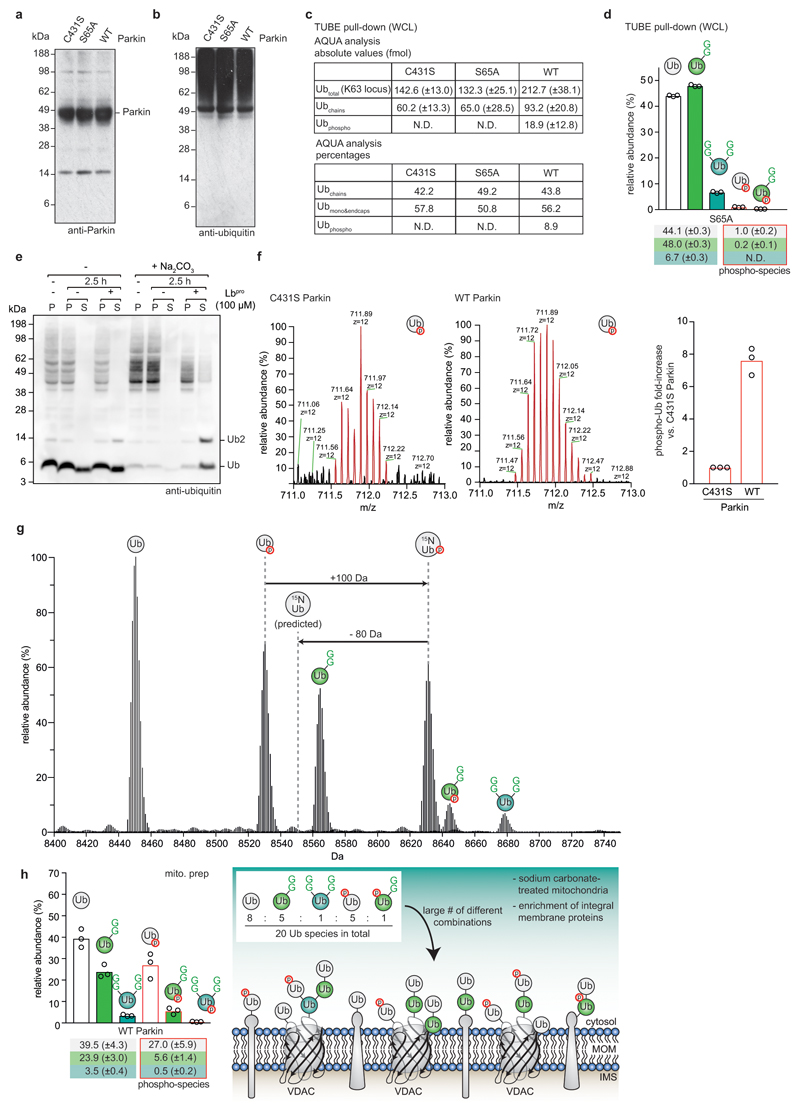
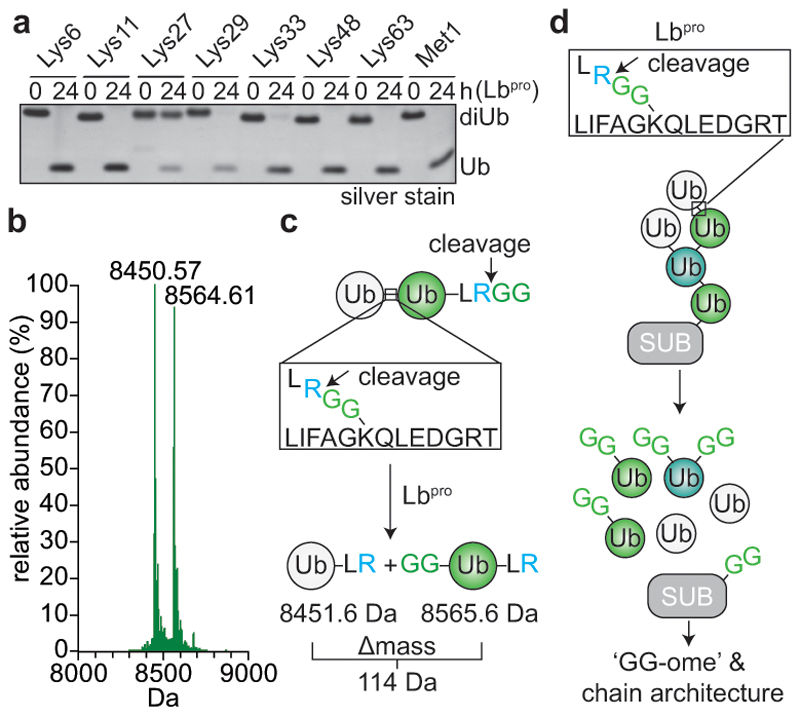
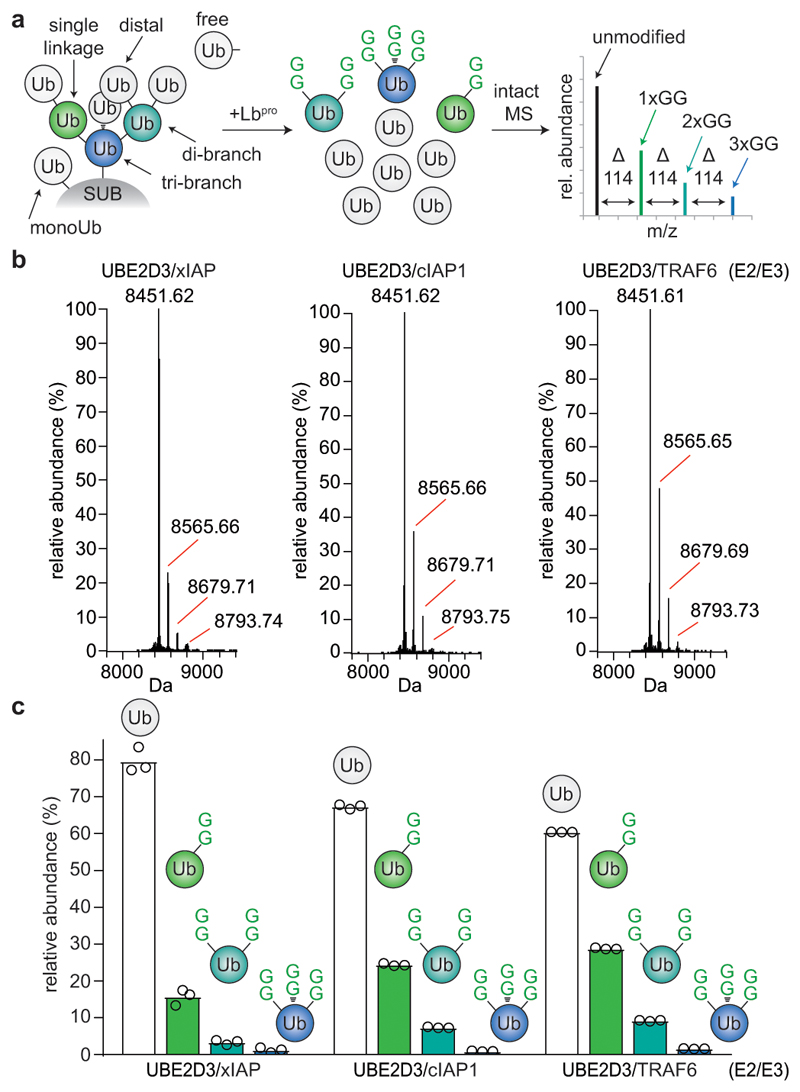
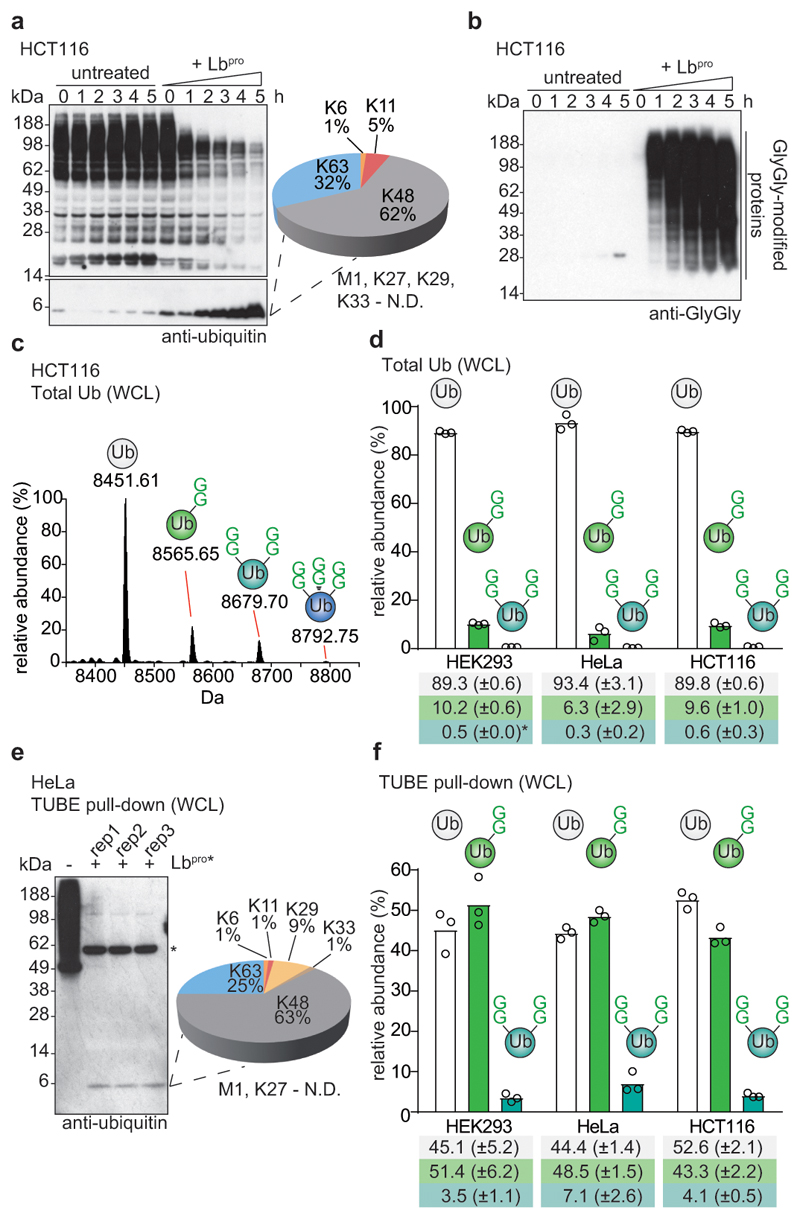
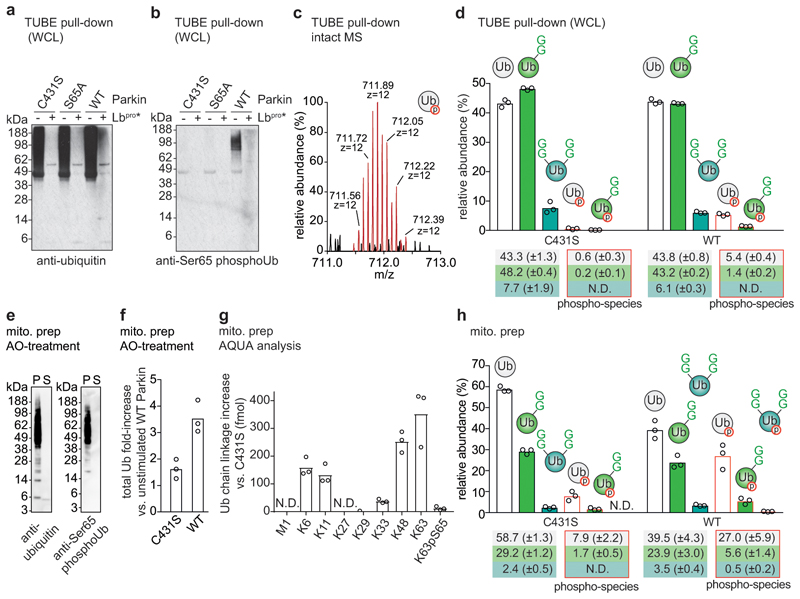
Protein ubiquitination is a multi-functional post-translational modification that affects all cellular processes. Its versatility arises from architecturally complex polyubiquitin chains, in which individual ubiquitin moieties may be ubiquitinated on one or multiple residues, and/or modified by phosphorylation and acetylation1-3. Advances in mass spectrometry have enabled the mapping of individual ubiquitin modifications that generate the ubiquitin code; however, the architecture of polyubiquitin signals has remained largely inaccessible. Here we introduce Ub-clipping as a methodology by which to understand polyubiquitin signals and architectures. Ub-clipping uses an engineered viral protease, Lbpro∗, to incompletely remove ubiquitin from substrates and leave the signature C-terminal GlyGly dipeptide attached to the modified residue; this simplifies the direct assessment of protein ubiquitination on substrates and within polyubiquitin. Monoubiquitin generated by Lbpro∗ retains GlyGly-modified residues, enabling the quantification of multiply GlyGly-modified branch-point ubiquitin. Notably, we find that a large amount (10-20%) of ubiquitin in polymers seems to exist as branched chains. Moreover, Ub-clipping enables the assessment of co-existing ubiquitin modifications. The analysis of depolarized mitochondria reveals that PINK1/parkin-mediated mitophagy predominantly exploits mono- and short-chain polyubiquitin, in which phosphorylated ubiquitin moieties are not further modified. Ub-clipping can therefore provide insight into the combinatorial complexity and architecture of the ubiquitin code.
Conflict of interest statement
The authors declare no competing interests.
Figures
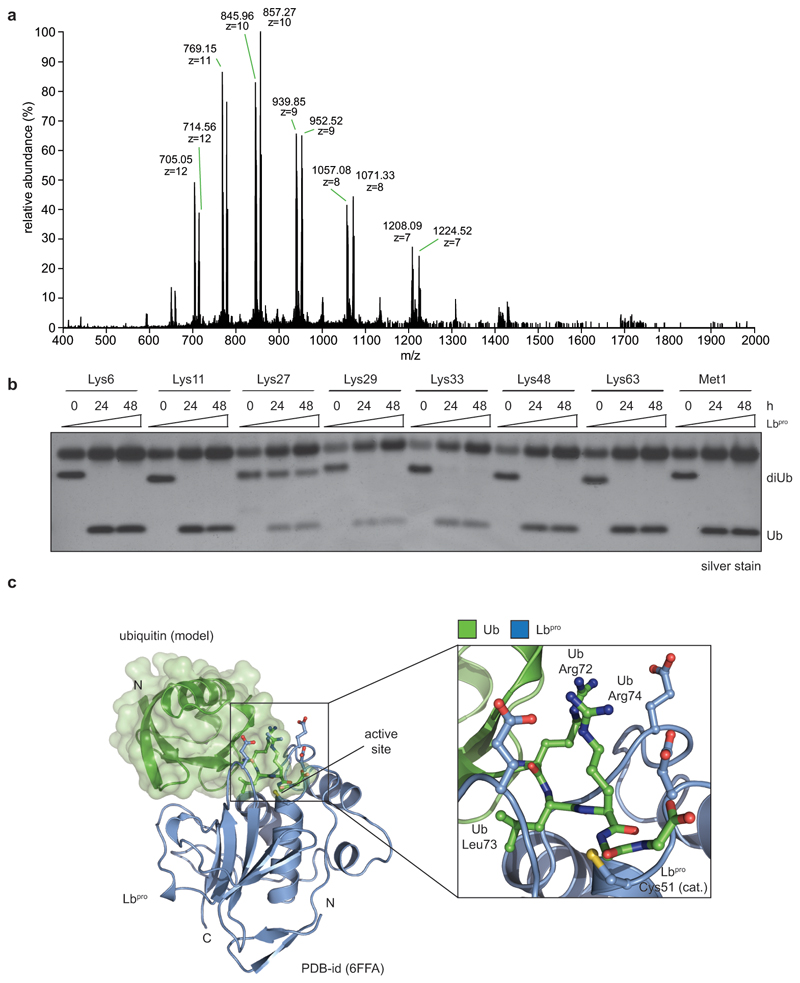
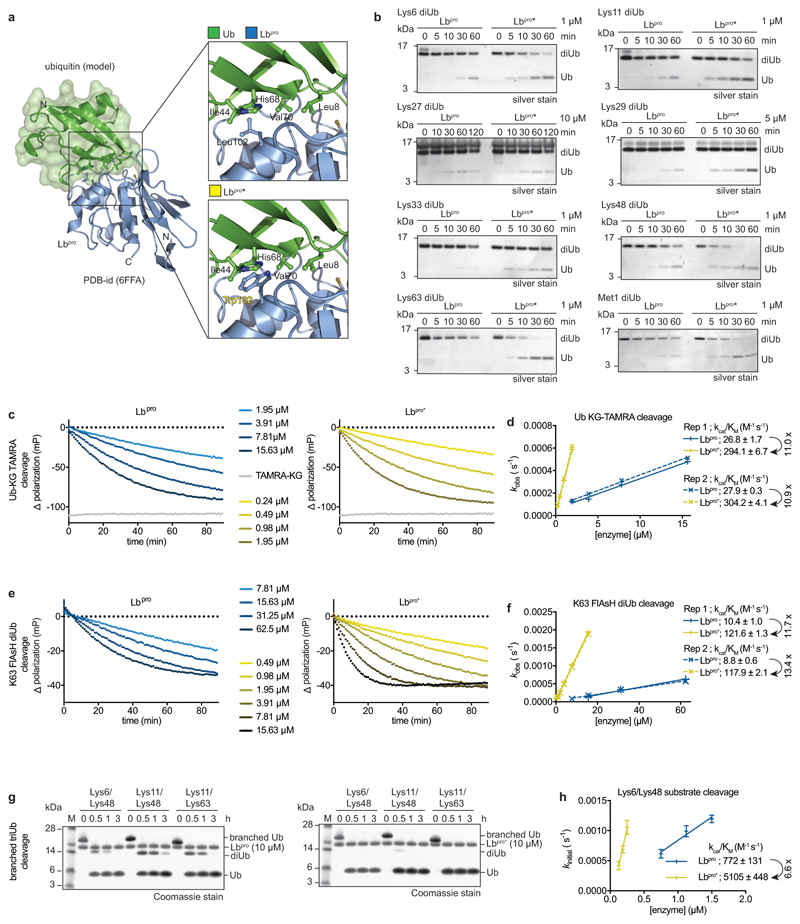
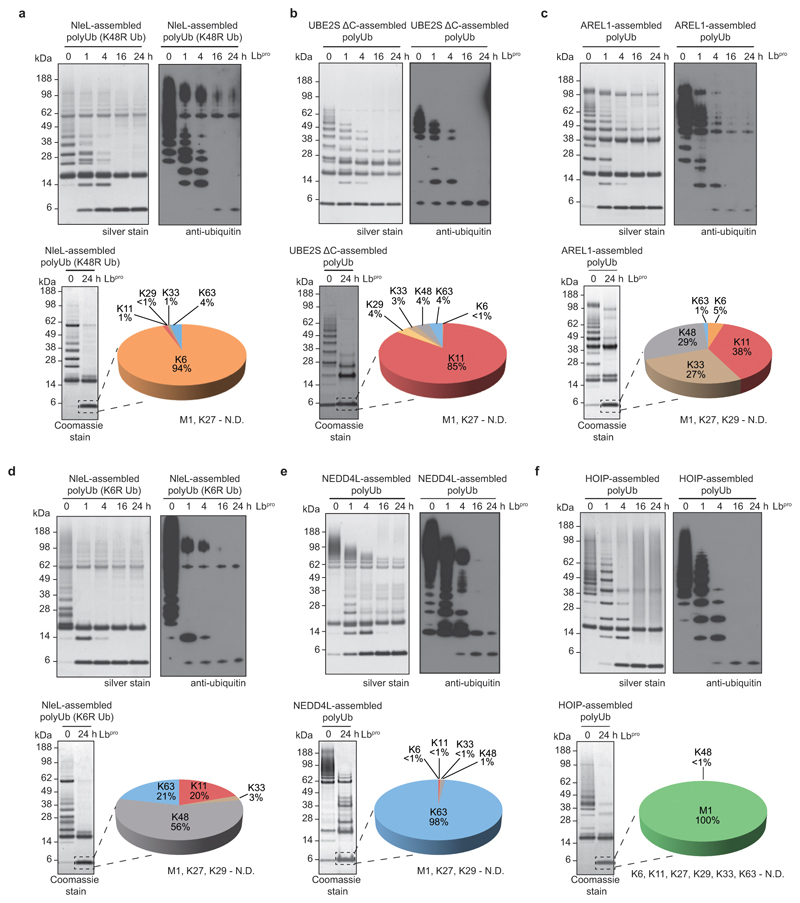
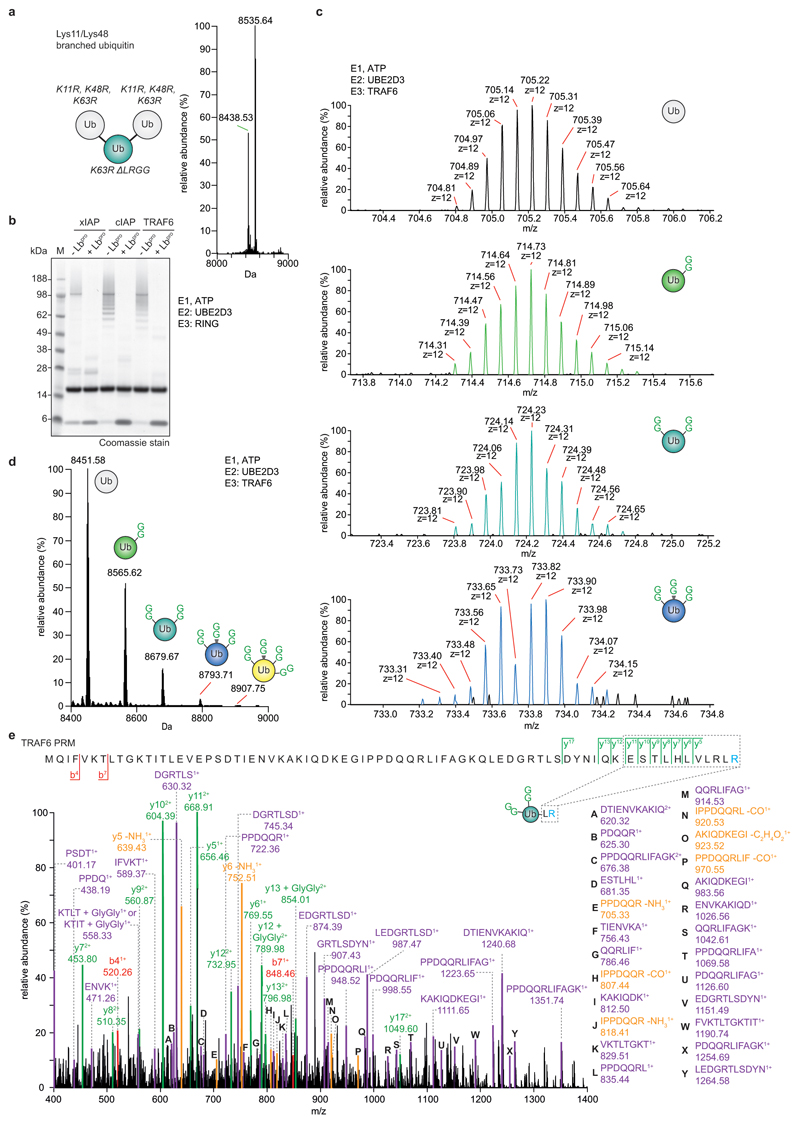
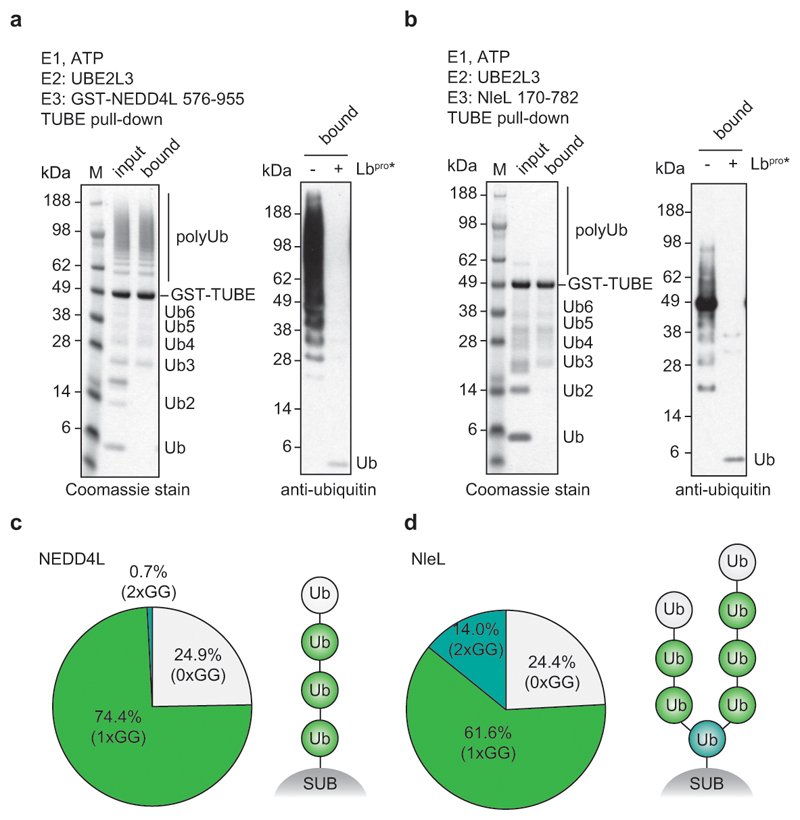
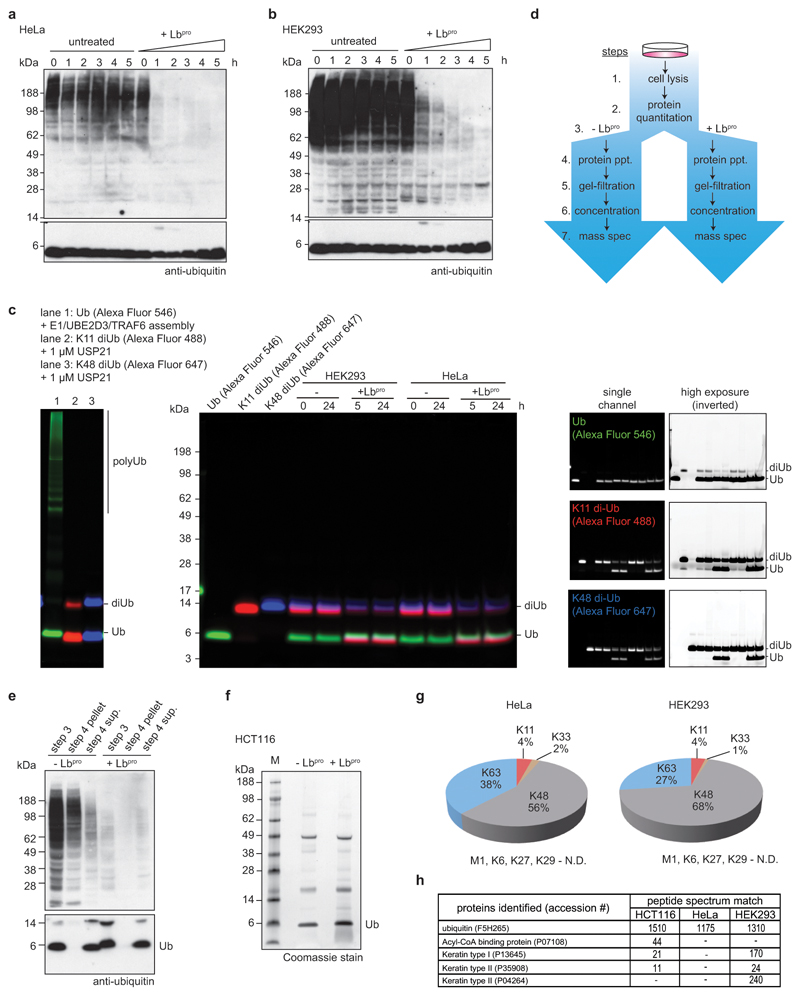
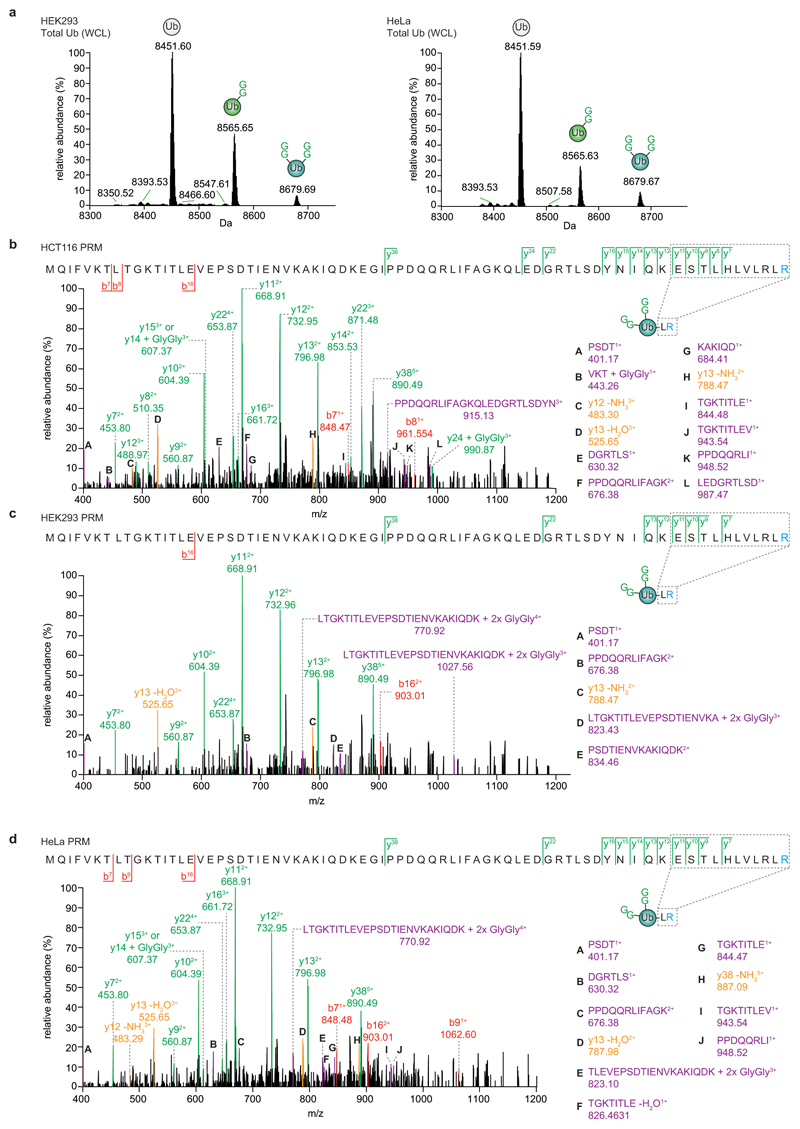
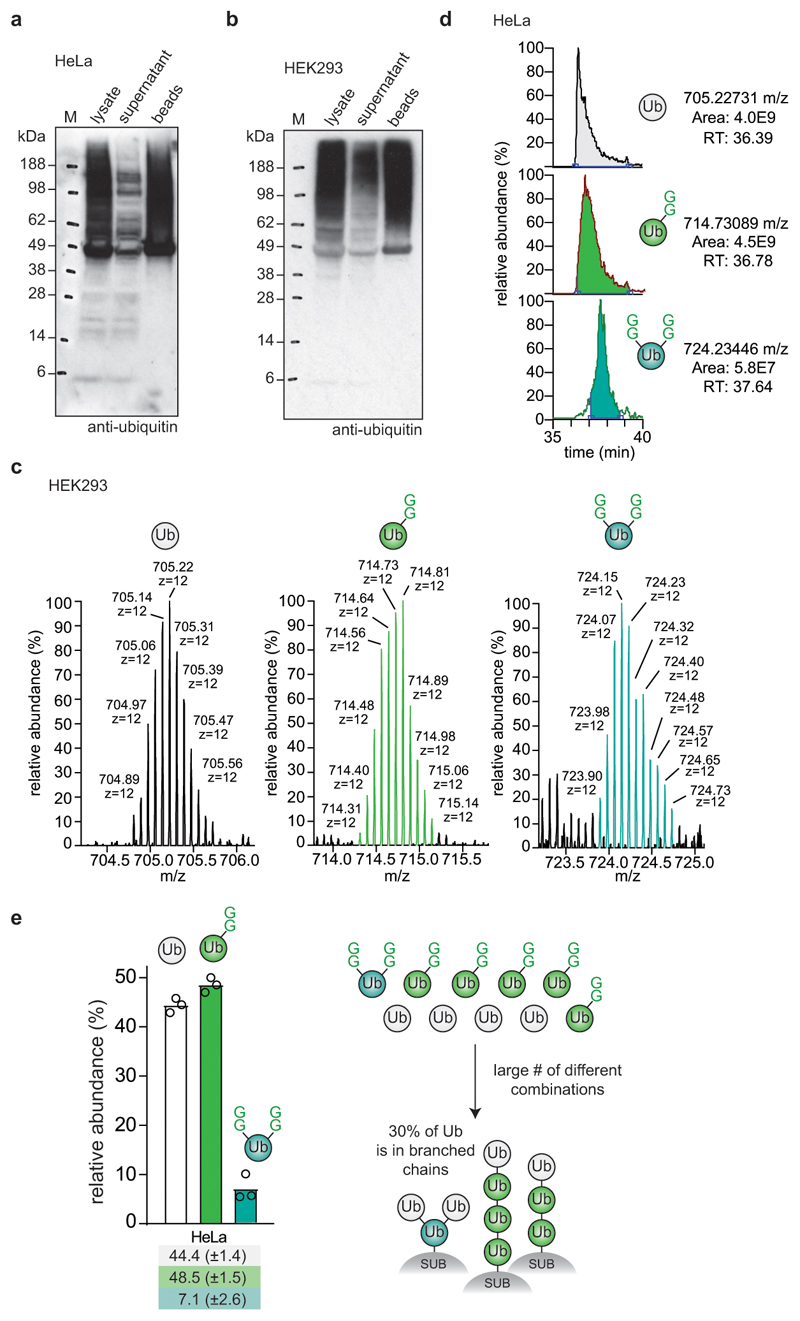
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
