

Regulation of mitochondrial metabolism in murine skeletal muscle by the medium-chain fatty acid receptor Gpr84
- PMID: 31415180
- PMCID: PMC9292139
- DOI: 10.1096/fj.201900234R
Regulation of mitochondrial metabolism in murine skeletal muscle by the medium-chain fatty acid receptor Gpr84
Abstract
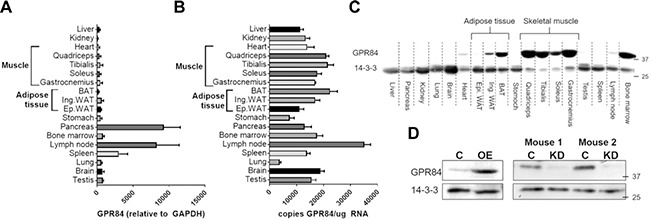
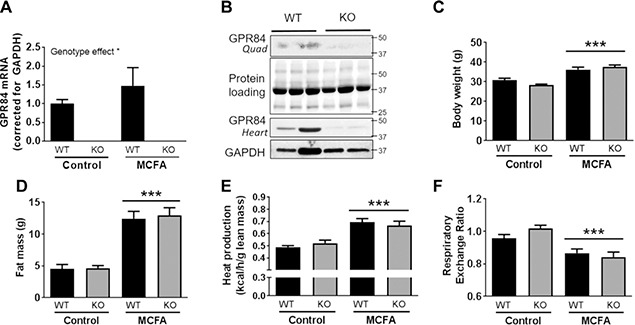
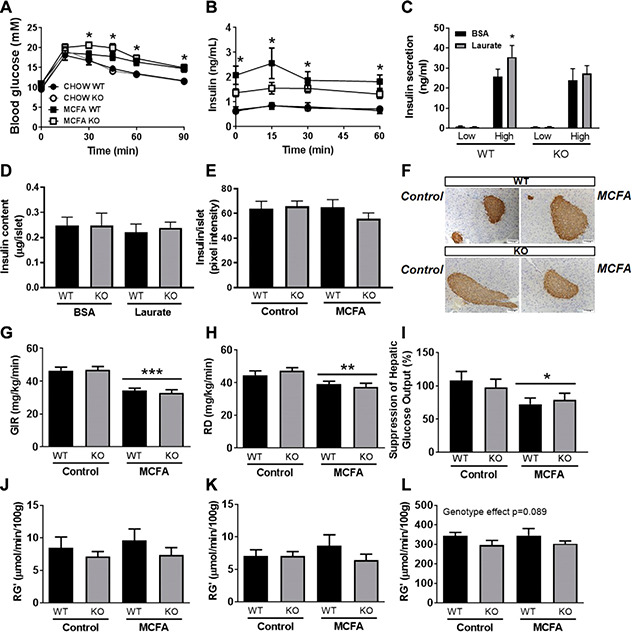
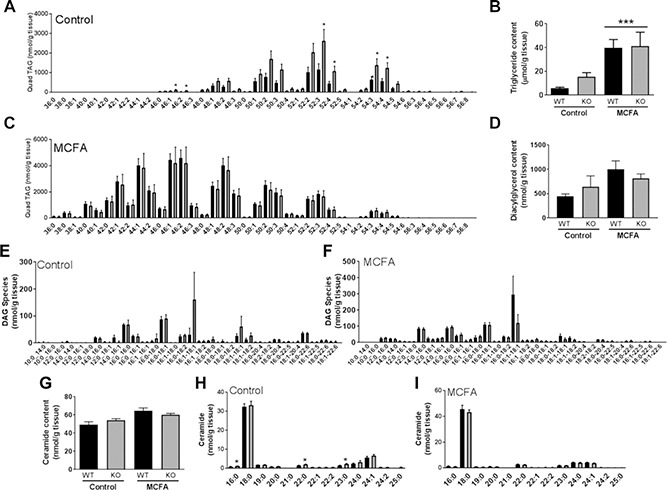
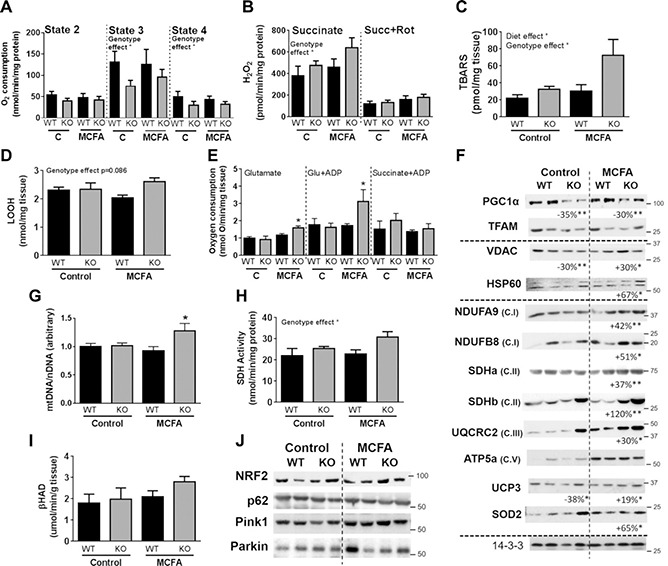
Fatty acid receptors have been recognized as important players in glycaemic control. This study is the first to describe a role for the medium-chain fatty acid (MCFA) receptor G-protein-coupled receptor (Gpr) 84 in skeletal muscle mitochondrial function and insulin secretion. We are able to show that Gpr84 is highly expressed in skeletal muscle and adipose tissue. Mice with global deletion of Gpr84 [Gpr84 knockout (KO)] exhibit a mild impairment in glucose tolerance when fed a MCFA-enriched diet. Studies in mice and pancreatic islets suggest that glucose intolerance is accompanied by a defect in insulin secretion. MCFA-fed KO mice also exhibit a significant impairment in the intrinsic respiratory capacity of their skeletal muscle mitochondria, but at the same time also exhibit a substantial increase in mitochondrial content. Changes in canonical pathways of mitochondrial biogenesis and turnover are unable to explain these mitochondrial differences. Our results show that Gpr84 plays a crucial role in regulating mitochondrial function and quality control.-Montgomery, M. K., Osborne, B., Brandon, A. E., O'Reilly, L., Fiveash, C. E., Brown, S. H. J., Wilkins, B. P., Samsudeen, A., Yu, J., Devanapalli, B., Hertzog, A., Tolun, A. A., Kavanagh, T., Cooper, A. A., Mitchell, T. W., Biden, T. J., Smith, N. J., Cooney, G. J., Turner, N. Regulation of mitochondrial metabolism in murine skeletal muscle by the medium-chain fatty acid receptor Gpr84.
Keywords: insulin resistance; insulin secretion; mitochondrial function.
Figures
References
-
- Kraegen, E. W. , and Cooney, G. J. (2008) Free fatty acids and skeletal muscle insulin resistance. Curr. Opin. Lipidol. 19, 235–241 - PubMed
-
- Elagizi, A. , Lavie, C. J. , Marshall, K. , Di Nicolantonio, J. J. , O'Keefe, J. H. , and Milani, R. V. (2018) Omega‐3 polyunsaturated fatty acids and cardiovascular health: a comprehensive review. Prog. Cardiovasc. Dis. 61, 76–85 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
