

Alternate subunit assembly diversifies the function of a bacterial toxin
- PMID: 31417089
- PMCID: PMC6695444
- DOI: 10.1038/s41467-019-11592-0
Alternate subunit assembly diversifies the function of a bacterial toxin
Abstract
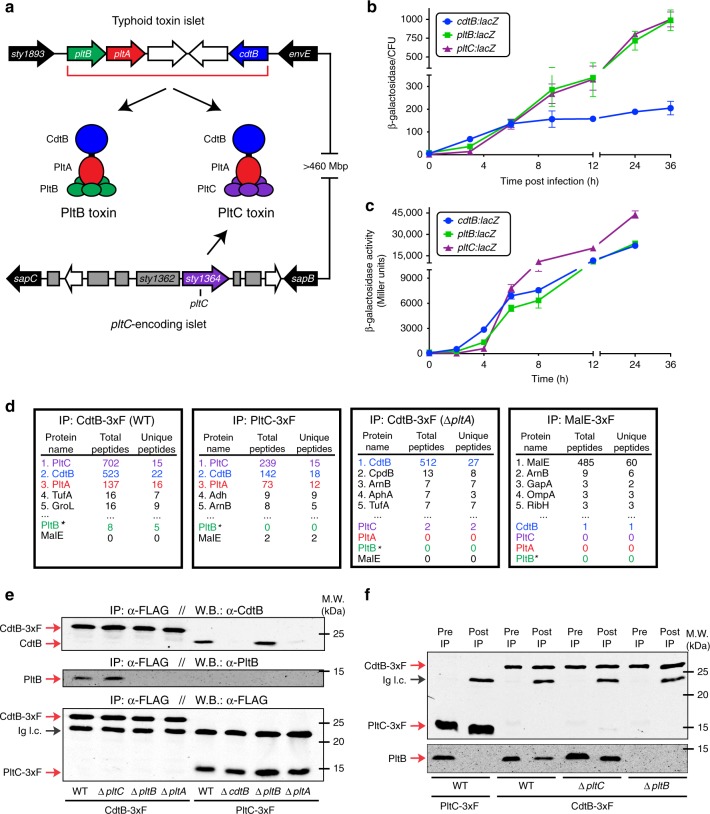
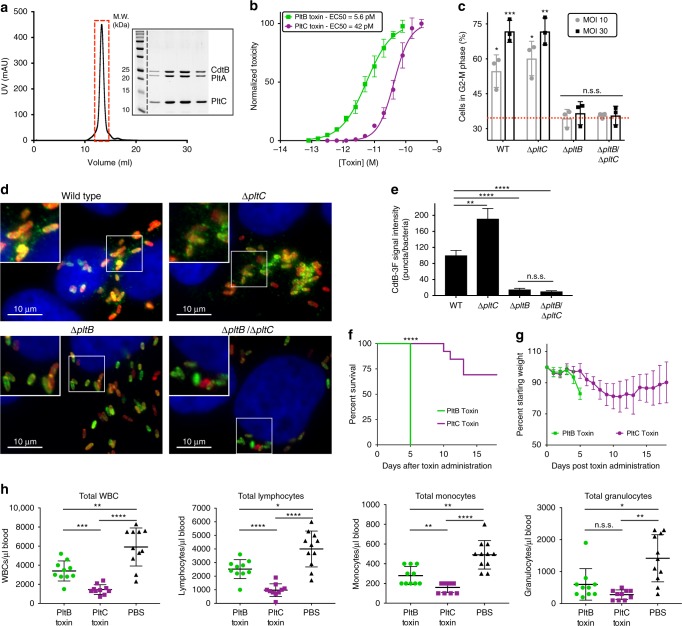
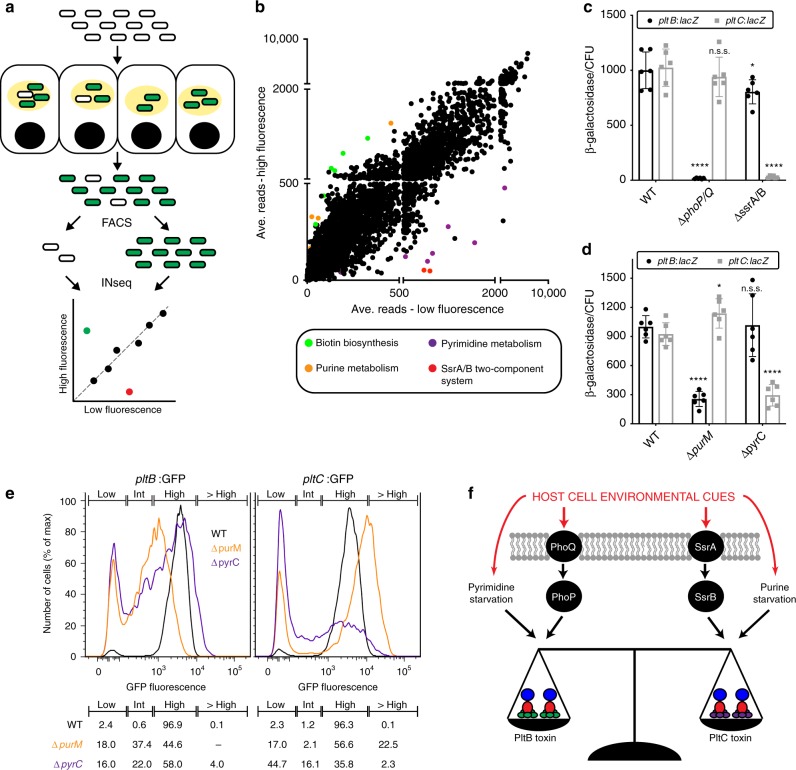
Bacterial toxins with an AB5 architecture consist of an active (A) subunit inserted into a ring-like platform comprised of five delivery (B) subunits. Salmonella Typhi, the cause of typhoid fever, produces an unusual A2B5 toxin known as typhoid toxin. Here, we report that upon infection of human cells, S. Typhi produces two forms of typhoid toxin that have distinct delivery components but share common active subunits. The two typhoid toxins exhibit different trafficking properties, elicit different effects when administered to laboratory animals, and are expressed using different regulatory mechanisms and in response to distinct metabolic cues. Collectively, these results indicate that the evolution of two typhoid toxin variants has conferred functional versatility to this virulence factor. More broadly, this study reveals a new paradigm in toxin biology and suggests that the evolutionary expansion of AB5 toxins was likely fueled by the plasticity inherent to their structural design coupled to the functional versatility afforded by the combination of homologous toxin components.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
